- ��x��(qu��n)��
- 200
- UID
- 935
- �e��
- 20185
- ���}
- 1143
- ����
- 5112
- ���A
- 21
- ��(j��ng)�(y��n)
- 20185 �c(di��n)
- ����
- 15855 Ԫ
- ע��(c��)�r(sh��)�g
- 2010-12-27
TA��ÿ������ | ����ȥ��
2014-10-22 08:04 |
|---|
�����씵(sh��): 17 �� [LV.4]ż������III - ����
- 5112
- ���A
- 21
- ��(j��ng)�(y��n)
- 20185 �c(di��n)
- ����
- 15855 Ԫ
- ע��(c��)�r(sh��)�g
- 2010-12-27

|
ˮ�a(ch��n)��(d��ng)��I(y��ng)�B(y��ng)�c����о�����
������
�B�T��W(xu��)�����c�h(hu��n)���W(xu��)Ժ
ˮ�a(ch��n)���ճ�τ�
1.ճ�Y(ji��)��
ճ�Y(ji��)��Ҳ�Q�x�΄�����Ҫ���ڼӹ��w����ϣ��Ը�������Ʒ�|(zh��)(�������ʡ�Ӳ�ȡ���ĥ��)���������a(ch��n)Ч�ʼ����L(zh��ng)�Tģ���������ɜp����Ϸۉm�������w���ɫ��(w��n)�����e��ˮ�a(ch��n)�~�r���M�����D����Ҫ�������ˮ�в�ɢ������������,�Ķ��p���B(y��ng)����ʧ���õ����ӄ������ɷ���Ȼͻ��W(xu��)�ϳ�ɴ��ˮ�a(ch��n)��ό�(du��)���ճ�τ���Ҫ���^�ߣ������(l��i)����(gu��)��(n��i)�⌦(du��)ˮ�a(ch��n)���ճ�τ����о��dz����S���µ�ճ�τ��N����Ƴ������Ƴ���ճ�Y(ji��)�������ϿɝM����N�B(y��ng)ֳ��(du��)�����Ҫ��
2.���ճ�Y(ji��)���ķ��
2.1��Ȼ�
(1)��ľ���ڵ��z֭:�����z�������z�����z�ȡ��@�ճ�Y(ji��)�����ܵ�pHֵ����ȡ��V���|(zh��)�}�����|(zh��)��Ӱ푶�����ճ�ȣ����δ�ܵõ��V����(y��ng)�á�
(2)ϡ����ճ����ճ�Y(ji��)��:����(r��n)�����������c���������ȡ��@Щճ���V��ճ���^�ͣ����ӱ�������(y��ng)�Ӵ���(j��)�Q�������_(d��)�V������о������Ƶ�202�����ճ�Y(ji��)�������H�m��������������Ҳ�������䉺������ ˮ�a(ch��n)���ճ�τ�
(3)ֲ�����:��С�������ס�ľ�����R����ȵ��ۻ�׃�Ե��ۡ��@�ճ�Y(ji��)��ճ�Y(ji��)���Ĵ�С��ȡ�Q�ڵ�����ͺ���ϼӹ��O(sh��)���Լ��������g(sh��)��
(4)������z�|(zh��):�к������c�������z����֬�ȡ������ճ�Y(ji��)��һ��Y(ji��)������(qi��ng)�����r(ji��)���F��һ������ԇ�(y��n)�о�����С�
2.2 �˹��ϳ��
(1)�u���w�S��:��Ʒ���ɫ�w�S����w�����ĩ���o(w��)�����o(w��)ζ���������ԣ�ˮ��Һ��(du��)���(w��n)������ճ���S�ضȻ�߶����͡���������������˳��^(gu��)2%��
(2)��ȩ��֬:��Ʒ�ڸ��N�䡢�ᡢ��ͻ��W(xu��)���g�ȿ��̵ĭh(hu��n)���������õķ�(w��n)���ԣ����Ҿ߂�r(ji��)�������������(y��u)�c(di��n)�����w�����ճ�τ����^�õĂ��x���ϡ���������0.5%�^����m��
(3)ľ�|(zh��)�ػ����}:��һ�N��������Ѿۺ������ɫ���̑B(t��i)�������ԏ�(qi��ng)���w������ϳ�Ʒ�к������ɳ��^(gu��)4%��
(4)�����І�λ��ԺУ�������(l��i)�����_�l(f��)�����ƏV��(y��ng)�õ����ճ�Y(ji��)������V��ԭҰ��˾�ľ����ߡ��������ճ�Y(ji��)���a(ch��n)Ʒ��
���⣬߀�о���ϩ��(PUA)���۱�ϩ�ᡢ���N���������}��ľ�|(zh��)�ء�HJ-l�������c���ۼ��塢�������c�c�����ȣ����������ճ�Y(ji��)����
ͬ�r(sh��)������(j��)ճ�Y(ji��)���Ƿ�(du��)�B(y��ng)ֳ��(d��ng)���РI(y��ng)�B(y��ng)���ã��ɷ֞�I(y��ng)�B(y��ng)���c�ǠI(y��ng)�B(y��ng)�̓ɴ��ǰ�����~�r�{��� ֲ����ۡ���Ȼ�V���|(zh��)�Լ�һЩ�˹��ϳɵ���N87��(���Cʡ�Zʳ�ƌW(xu��)�о�������)ճ�Y(ji��)���ȡ�����ָ��(du��)��(d��ng)��o(w��)�I(y��ng)�B(y��ng)�r(ji��)ֵ��ճ�Y(ji��)�������ľ�z�����w�S�صȡ�
3 ���ճ�Y(ji��)����(y��ng)�߂�����c(di��n)
3.1 ��(du��)����и��N�I(y��ng)�B(y��ng)�ɷ־��������ճ�϶ȣ����C�I(y��ng)�B(y��ng)ȫ�r(ji��)���ܷ�ֹɢʧ��Ⱦ��������\(y��n)ݔ�^(gu��)���в������飬�ۉm�٣�
3.2 �������a(ch��n)��ȡ�����й��I(y��)�����a(ch��n)�Ŀ����ԣ�������Þ��(d��ng)��ĠI(y��ng)�B(y��ng)�أ�����(du��)��(d��ng)����������ա���(n��i)������������(j��ng)���������⡢���L(zh��ng)���wɫ�ȟo(w��)����Ӱ푣�
3.3 �����^�ߵĻ��W(xu��)��(w��n)���Ժ᷀͟(w��n)���ԣ����c������ϳɷְl(f��)���������W(xu��)����(y��ng)��
3.4 �o(w��)�����o(w��)������ζ�����������m���Ժ��Tʳ�ԣ�
3.5 �����٣���ϣ��ɱ��ͣ�����(l��i)�B(y��ng)ֳ��(j��ng)��(j��)Ч�棻
3.6 ��(du��)�A(y��)���ϻ������ϵļӹ�����ˇ�]���e���̵�Ҫ��
3.7 �������õı����ԣ��Ա��C����ˮ�в���ɢʧ��
3.8 ���õ�ճ���Ժͳ�ˮ�ԣ�����ճ�τ�����Ѹ���γ�ճ���Ժ�(qi��ng)�������w������ˮ�֝B����ό�(sh��)�w����������ϵ���(r��n)�����͡�
4.�x��ճ�τ���(y��ng)���]�Ďׂ�(g��)���棺
4.1�B(y��ng)ֳ��(d��ng)���ʳ�Լ��䌦(du��)�D����ˮ�з�(w��n)���Ե�Ҫ�� һ���ǔzʳ�^����ˮ�a(ch��n)��(d��ng)��Ҫ���^�õ�ˮ��(w��n)���ԣ����zʳ�^���ˮ�a(ch��n)��(d��ng)��t���^�͵�ˮ��(w��n)���ԣ�
4.2ճ�τ������|(zh��)���m�����ͳɱ���
4.3ճ�τ��c��ϳɷֵ�����ã������Ƿ��(hu��)�ƉĠI(y��ng)�B(y��ng)�ɷ֣����}�����ߕr(sh��)�������z��ճ���½������^�ۺ������ӕr(sh��)�������z������ҲҪ���ӣ��@�c�����z���}�x�Ӯa(ch��n)������������ճ�����P(gu��n)��
4.4ճ�τ��ĠI(y��ng)�B(y��ng)�r(ji��)ֵ��ճ�τ�������͵����|(zh��)����ṩһ���ĠI(y��ng)�B(y��ng)������߀Ҫ���]ճ�τ���(du��)ˮ�a(ch��n)��(d��ng)�����L(zh��ng)�l(f��)���ʹ���Ӱ푣�������^(gu��)����(hu��)Ӱ��~����L(zh��ng)��
4.5 ����ճ�τ����_��ʹ�÷��������ĕr(sh��)�g���ضȺ�ˮ�֣�ԭ�����r��������l�����ã���(y��ng)���ڸ��ꎛ�̎����?y��n)�ճ�τ������?hu��)�l(f��)���Խ�(li��n)�����Խ�ߣ�ԓ����(y��ng)Խ�죬
���⣬ճ�τ��a(ch��n)Ʒ����������������t�Y(ji��)�ɿ��ʧȥ�Y(ji��)���ܡ�
5. ��ˮ�a(ch��n)�B(y��ng)ֳ�I(y��)�ϵđ�(y��ng)��:
(1)��(j��)�ۼt�ǵ�(1999)�о��Y(ji��)��������������m����ϡ������������~����ø��֬��ø�͵���ø�Ļ��ԣ����M(j��n)�~�w��(du��)�I(y��ng)�B(y��ng)���|(zh��)���������գ����ܴ��M(j��n)���L(zh��ng)�������~���Ʒ�|(zh��)��������Ȫ��1997�������з�ӳ:ʹ��ճ�������ճ�Y(ji��)�������L(zh��ng)ˮ�a(ch��n)�����ˮ�����ݕr(sh��)�g���Ұ��\(y��n)�r(sh��)�����Ɠp������(du��)�A(y��)���~��������ˮ�|(zh��)���зe�O���á�
(2)��(j��)���J��(1996)��(b��o)��:���ճ�Y(ji��)��N87��ʹ�����漰����N��ˮ��ˮ�~��������N�~�r�����~�����~�����V���Oؐ����ؐ���з�ȡ����ң��ڹ�ˇ�ϣ���B(y��ng)�ж��յ�
�˺ܺõ�Ч����
(3)�����A(1996)�J(r��n)�飬��-������Ŀǰճ�Y(ji��)���еļ�Ʒ�����~��(du��)��-���۵�����Ч����á�ͬ�r(sh��)���w���⣨200O���J(r��n)�飬���~����Ц�-���۵ı��������^(gu��)18%��ã�����Ӱ푼��~�������L(zh��ng)��
(4)��(j��)�O��|(1996)��B��ϡ���ܴ̼��~�ʳ��������(qi��ng)�������ܣ����μ��������Kˮ�a(ch��n)�о�����ϡ���c�ך��ط���(y��ng)�Ƴ���(du��)�r�D��ճ�Y(ji��)��������3%~5%����߮a(ch��n)��20%���ҡ����������c�����r�Ͽ����L(zh��ng)�����ˮ�зֽ�r(sh��)�g���̼��~�rʳ�����{(di��o)��(ji��)�C(j��)�ƴ��x����ֹˮ�|(zh��)��Ⱦ��
6.���ճ�Y(ji��)���ڑ�(y��ng)���Б�(y��ng)ע��Ďׂ�(g��)��(w��n)�}
6.1 ���ӕr(sh��)��(y��ng)����(j��)�ɱ���Ŀ�ġ�Ч���x��������m��ճ�Y(ji��)����
6.2 ճ�Y(ji��)����ʹ�Ä�(sh��)�����ӳɱ����ڟo(w��)��(j��ng)��(j��)Ч��r(sh��)��(y��ng)�M������ʹ�á�
6.3 ����(j��)�B(y��ng)ֳ��(du��)��ͬ���x��ͬ���ܵ�ճ�Y(ji��)���������õ�������
6.4 ճ�Y(ji��)���a(ch��n)Ʒ���x��M���ܵذ���(gu��)�Ҙ�(bi��o)��(zh��n)��(zh��)�С�
�C�������������a(ch��n)��(sh��)�`�У���(y��ng)����(j��)����������a(ch��n)���c(di��n)���B(y��ng)ֳ���a(ch��n)�^(gu��)���е������ԣ����x���ʹ�þC�����ܱ��^��(y��u)Խ��ճ�Y(ji��)��,���_(d��)����߽�(j��ng)��(j��)Ч���Ŀ�ġ�
7.ˮ�a(ch��n)���ճ�τ�ճ�������u(p��ng)�r(ji��)ָ��(bi��o)
A.������
��ͬ���B(y��ng)ֳ�N����L(zh��ng)�ں�ʳ�Ե��~���r��ˮ�a(ch��n)��(d��ng)�(du��)����ΑB(t��i)��Ҫ���ͬ���鱣���@һ�ΑB(t��i) ,��Ҫ����������õı����� ,�Ա��C����ˮ�в���ɢʧ����(du��)�� ,���ڼӹ���ˇ�ϲ�ȡijЩ��ʩ�� (�����g(sh��)�y�ȴ��O(sh��)��Ͷ�Y����ϳɱ��� ) ,ͨ�����ü���ճ�τ��ķ������� :ܛ�w����ό�(du��)�����~��^���m ,Ӳ�w����ό�(du��)���~�����~�����~���F(tu��n)�^�����~��^�m�� ;�B(y��ng)���ú�������^�� ,���ɷ��ϰ������ ,��ˮ��ճ�τ����{(di��o)�Ƴ� ,���Џ��ԡ�ճ�ԡ���չ�� ,��ˮ�в�����ɢ ;�r��з�ȼך�Ĕzʳ���c(di��n)�� ,ֻ������ˮ�ĉK����� ,�������ĉK������ϵ���Ϣ̎�� ,�L(zh��ng)�r(sh��)�g�����zʳ ,�����ϵ��ΑB(t��i)����m���r��зĔzʳ��(x��)�ԡ�
B.ճ�����ܣ��w����ȫɢ�_�ĕr(sh��)�g����
C.ճ�τ��ﻯ���ܜy(c��)��
a.������:�Qȡճ�τ�20�磬һ������ʢ��50mlˮ�ğ����У��^���ճ�τ���ˮ�н���(r��n)��min��30min�Լ�1h��2h��6h��12h��24h��������B(t��i)��
b.�Ҹ���: ���b�Mˮ��100ml��Ͳ�y(c��)ճ�τ��z�F(tu��n)�ij����ٶȡ�
c.��ˮ�ʣ�(ml ��g-1)
�Qȡճ�τ�20�磬һ������50mlˮ�ğ����У���ȫ������(r��n)���o��24h���ÞV���^(gu��)�V30min���ռ��VҺ������ˮ�ʰ���ʽӋ(j��)�㣺
��ˮ�ʣ�(50-�ռ��VҺ)/ճ�τ�ԇ����
8.ˮ�a(ch��n)���ճ�τ�ճ�������u(p��ng)�r(ji��)
A.ճ�����ܣ�ճ�τ���ͬ���ӱ���ԇ�(y��n)�c�D�ϲ��ӟᡢ�ӟἰ�����ճ������ԇ�(y��n)������������ճ�τ��ij����D�ϼӟ��ճ�����ܾ��@����ߣ��@�c�D���Ц�- ���۵ĺ������P(gu��n)��ճ�τ������������D��ճ�Y(ji��)�ԣ������Ƕ��r(ji��)���ᣨ���ᡢ�����ᡢ��ʯ�ᣩ��(qi��ng)���~���c��- ���۵�ճ�Y(ji��)����
B.ճ�τ��ﻯ�����u(p��ng)�r(ji��)
ճ�τ��������ԡ��Ҹ��ԡ���ˮ�Ե��ﻯ���ܿ������u(p��ng)��ճ�τ���(y��u)�ӵ�����(j��)֮һ��һ����r���õ�ճ�τ����ܻ�����ˮ�������^��(qi��ng)�������ԣ���ˮ����һ����ˮ�ʺ��ԣ���̎���^��(w��n)����������B(t��i)��ԇ�(y��n)������ħ�ۡ��������c�������ﻯ�����^��(y��u)��ħ�ۡ�������ˮ����Ѹ�ٽ���(r��n)�γ��z�w���������c����(r��n)�^�������������ܮa(ch��n)���^��(qi��ng)���������ҳ�ˮ�ʸߡ�
C. Ӱ������ˮ�Ե�����
��1��ԭ�ϵĽM�ɼ������
ԭ�ϵĽM�ɼ�����Ȍ�(du��)�����ˮ�Ե�Ӱ��^��һЩԭ�Ͽ���������ˮ�� ,����?c��)�����䷽����ռ�ı����?,�a(ch��n)Ʒ����ˮ�Ծͺá��෴ ,����ʹ����һЩԭ�� ,�a(ch��n)Ʒ����ˮ�Ծ��п���׃�ϵ�y(t��ng)����ˮ�a(ch��n)��ϳ��õ�ԭ�� ,�l(f��)�F(xi��n)һЩԭ�ό�(du��)�a(ch��n)Ʒ��ˮ�Ե���ؓ(f��)Ӱ푴��w���О� :��ۡ������ɡ�С�����~�ۡ������ɡ����ɡ��QӼ����Ƥ�������S�ۡ����ס�����ˮ�a(ch��n)��ϵĵ��������@����������ϡ����~�ۡ���ɵ���Ҫ����ԭ�ϵ�ճ�Y(ji��)�Զ�������ۡ����a(ch��n)�ϳ����õ��� (��(j��ng)����̎�� )�ĸ�ճ�Y(ji��)�ԁ�(l��i)��������w������ˮ�ԡ����� ,߀�������������ճ�Y(ji��)�� ,�Ը������Ʒ�|(zh��)��
��2��ԭ������ (���� )
ԭ�����ȼ�(x��)�t����e���^�� ,�ɫ@���^�õ��{(di��o)�|(zh��)Ч�� ,�w����ճ�Y(ji��)�Ծͺ� ,Ӳ���^�� ,ˮ�з�(w��n)��������(qi��ng) ;ԭ�������^(gu��)�� ,�Ƴ��w�����^������ ,��(w��n)�����^�� ������ ,�^(gu��)�ȷ����(hu��)
ʹ���a(ch��n)�������� ,�������� ,���a(ch��n)�ɱ����ӡ���� ,�����a(ch��n)�^(gu��)���Б�(y��ng)����(j��)�w���ϵ�Ҫ���x���m�˵ķ������ȡ��~��������(ji��n)���Ҷ� ,����ø���w�صͶ����Բ��� ,��(du��)���ԭ�ϵķ�������Ҫ���^��(y��n)��
ˮ�a(ch��n)���ճ�τ�
��3���{(di��o)�|(zh��)
��ϏS�ձ���������{(di��o)�|(zh��) ,��ֱ�ӌ�����ͨ�������M(j��n)��ˮ��̎�����r(sh��)�g���ضȡ�ˮ�����{(di��o)�|(zh��)����Ҫ�ء�
3 . 1���{(di��o)�|(zh��)�r(sh��)�g
��һ���ķ�����(n��i) ,�{(di��o)�|(zh��)�r(sh��)�gԽ�L(zh��ng) ,ԭ�ϵ��컯��Խ�� ,���ۺ�����Խ�� ,ճ�Y(ji��)�Ծ� ,����Ч���� ,����|(zh��)���á���֮ ,����|(zh��)���ˮ�a(ch��n)���Ҫ�����^�L(zh��ng)����ˮ�r(sh��)�g ,��(du��)�w�����컯�ȡ�ճ�Y(ji��)��Ҫ���^�� ,���^�L(zh��ng)���{(di��o)�|(zh��)�r(sh��)�g ,���a(ch��n)�Ͽɲ��ö���{(di��o)�|(zh��)�����p�S�{�~�{(di��o)�|(zh��)���ȁ�(l��i)�����{(di��o)�|(zh��)�r(sh��)�g��
3.2�{(di��o)�|(zh��)�ض�
�{(di��o)�|(zh��)�ضȌ�(du��)�w����ˮ�Ե�Ӱ��^���{(di��o)�|(zh��)�ض�Խ�� ,��ˮ��Խ�� ,���ض�̫�� (100������ )��ʹ���ϝ���^(gu��)�� ,��(d��o)��ģ���� ,�������a(ch��n)�����w�����ɫƫ�� ,Ӱ푮a(ch��n)Ʒ�����^ ;�{(di��o)�|(zh��)�ضȵ� (80������ ) ,��ˮ���^�� ,ͬ�r(sh��)Ҳ��(hu��)������������ġ��ڌ�(sh��)�H���a(ch��n)�� ,��(y��ng)����(j��)�䷽��ԭ�ϵ����Լ��a(ch��n)Ʒ���|(zh��)��Ҫ���x���{(di��o)�|(zh��)�ضȡ���ߵ����|(zh��)�c���w�S������䷽ (25%�� 45% ) ,�{(di��o)�|(zh��)�ض�ͨ���� 60�� 80�档�~�ϵ��{(di��o)�|(zh��)�ض�һ��Ҫ�� 85�����ϡ�
3.3��Ϸ���ˮ�ֺ���
ˮ�־��Н�(r��n)����������֮���á�Ҫ���Ƹ��|(zh��)�����w����� ,����Ҫ���m����ģˮ�� ,����Ҫ�m�˵Ĝضȡ���ģˮ����Ҫ��ԭ�ϱ�����ˮ�����{(di��o)�|(zh��)�^(gu��)�������������ˮ�ĺ�����ͨ�^(gu��)���ƻ�ϙC(j��)�л�Ϸ��ϵ�ˮ���Լ��{(di��o)�|(zh��)�r(sh��)���������ӿ��@���غͳ��m(x��)�ظ��ƽK�a(ch��n)Ʒ�w���ϵ��;öȡ�
�о��l(f��)�F(xi��n) ,��Ϸ��ϵ�ˮ�ֺ�����(du��)�w���;öȵ�Ӱ푘O�@������Ϸ��ϵ�ˮ��Խ�� ,�w������ˮ��Խ�� ,����ˮ��̫�� (�� 2 0�����ϕr(sh��) ) ,�����a(ch��n)�^(gu��)����������������ģ���� ,Ҳ��(hu��)ʹ�a(ch��n)Ʒ�y�Ը��
3.4 �M(j��n)���ٶ� (�a(ch��n)�� )
�M(j��n)���ٶȵĿ��� ,��(du��)�w������ˮ���ИO���Ӱ푡��M(j��n)���ٶȿ� ,�a(ch��n)���� ,��ԭ���е��ۼӟ�����r(sh��)�g�� ,��ʹճ�Y(ji��)���½� ,��ˮ�Բ� ;�p���M(j��n)���ٶ� ,���L(zh��ng)���ϼӟ�r(sh��)�g ,ʹ���۳�ֺ��� ,�w���ϵ���ˮ����� ,���a(ch��n)������ ,Ӱ����a(ch��n)Ч�ʡ���� ,��(y��ng)����(j��)�~���a(ch��n)�����w�������Ͳ�ͬ�������|(zh��)�� ,�����x���m�˵��M(j��n)���ٶȡ�
��4������
�҇�(gu��)Ŀǰʹ������ˮ�a(ch��n)�����ʽ�Dz��íh(hu��n)ģ�����C(j��)��ƽģ�����C(j��)���a(ch��n)����ͨӲ�w����ϡ��h(hu��n)ģ�Ŀ����s��(��ģ���c�h(hu��n)ģ����Ч��ȱ� )�c�w���|(zh��)�����P(gu��n)�������ԭ�ϽM�ɼ���ȡ����a(ch��n)��ˇһ����ǰ���� ,�h(hu��n)ģ�����s��Խ�� (1��1 7) ,�����ˮ�r(sh��)�gԽ�L(zh��ng) ;�h(hu��n)ģ�����s��ԽС(1��8) ,�����ˮ�r(sh��)�gԽ�� ����� ,Ҫ�����x��h(hu��n)ģ ,��ע���{(di��o)����ģ݁�g϶���е�λ�ü����_�ذ����E���������C(j��)�ȡ��е���ϏS�ڹ�ˇ�ϲ����؏�(f��)������(l��i)�����ϵ���ˮ�ԡ�
���������ӹ��������a(ch��n)��ˮ�a(ch��n)�w����ϱ��^�m���ڳ��Լ��ח������ˮ����(d��ng)�ˮ�a(ch��n)��ϳ��������ͻ�A���� ,�w�ϴ�С (���m�˵��w��ֱ�� )���~�w��С���P(gu��n)���~�w�� ,�����������(du��)��Щ������������~�������~ ,���w�����ӎ��f(w��n)�����ʮ�f(w��n)������� ,ᘌ�(du��)�~�IJ�ͬ���L(zh��ng)�� ,���������ܴ��������~���_����������H 50 �̣����� ,�����~������������� 4�� 6���� ,��������ϰٱ�����(gu��)��(n��i)���a(ch��n)С������ϵij��÷����������a(ch��n)�����^����w����� ,Ȼ�������鵽����ijߴ硣�������^(gu��)���� ,�o�ܵ��w������ӱ��Ɖ� ,���w����ˮ�����ڝ�ɢ ,�Ķ�Ӱ������w����ˮ�еķ�(w��n)���ԡ�
�~�N��ͬ,�䗫Ϣˮ�ӡ���ʳ�О�Ҳ��ͬ����Щ�~�ʳ�^��Ҏ(gu��)��,�����ܶ��r(sh��)�ص����Ͷι�c(di��n)��(l��i)������ʳ��ÿ��Ͷ� 10min���Ҽ��ܲ�ʳ�ꮅ���@�N�~�������ˮ�ԕr(sh��)�g�͟o(w��)����L(zh��ng) ,�����ˮ���ܾS��0 .5�����Ҽ��ɝM��Ҫ����Щ�~��ʳҎ(gu��)�����^�� ,������һ���ˮ����һ� ,Ȼ���ف�(l��i)��һꇡ������ʮ��犲��ܳ�ʳ�ꮅ���@Щ�~����Ͼͱ�����^�L(zh��ng)����ˮ�r(sh��)�g���r��M(j��n)ʳ�r(sh��)�����~��ǘ�һ�ڌ����w�������ǃ�(n��i),������ǰ�㡰�������w��������ʳ ,�����r�һ�w���߀δ����,���ӵ�ȥ����һ�w��ᘌ�(du��)�@�N��ʳ��ʽ,��ϵ���ˮ�r(sh��)�g������L(zh��ng)�����a(ch��n)��,�ɲ��ÔD�����g(sh��)���a(ch��n)���Եġ����ԵĻ��������
�~��� ,�ԝM�������B(y��ng)ֳģʽ�µĸ���(g��)�B(y��ng)ֳƷ�N�~���Ҫ��
(5)��s
��s�ǽ�����������w���ضȺ͝�ȡ�Ҫע���x���m�˵���s�r(sh��)�g����s�L(f��ng)�� ,�����w����Ӳ�Ⱥ���ĥ���ܵ�Ӱ푡�
ʹ����ɫ���ı�������ɫ���ĸ��
ˮ�a(ch��n)�B(y��ng)ֳ�ɂ��y(t��ng)�Ĵַ��B(y��ng)ֳ�l(f��)չ���F(xi��n)���ļ��s���S�����ܶ��B(y��ng)ֳ������ʹ���˹������ϣ��~���r��з���B(y��ng)ֳ���ڿs�̣�ֻ�ܫ@����������Ȼˮ��������������ЧɫԴ
���ҳɷֲ���(w��n)������(d��o)���~���r��з���wɫ׃�������ڴ��S�~���B(y��ng)ֳ�^(gu��)���У����S�~���wɫ��ʧȥ��ԭ�ȵĽ��Sɫ���w��׃���y����ƷζҲ�o(w��)���cҰ�������������y��Ӱ푵���Ʒ�ăr(ji��)�����⣬�^�p�~���wɫҲ�O���˂�����ҕ��ͨ�^(gu��)�����������һ�������|(zh��)�����ԏ��a(b��)�������㡣��ˣ���(du��)��ɫ��(��ɫ��)���о���ʹ���ѳɞ�F(xi��n)��ˮ�a(ch��n)�B(y��ng)ֳ�I(y��)����Ҫ��ʩ֮һ��
��ɫ����ָ����������ӣ������B(y��ng)ֳ��(d��ng)�����|(zh��)�����ɫ�ɵ����ӄ������ɷ���Ȼ���˹��ϳɃɴ��
�r�ۡ��S���ס��G��ȶ������õ�ɫԴԭ�ϣ�����ȻɫԴ�ɷֲ���(w��n)�������еĸ߃r(ji��)���ʱ���_�l(f��)��ɫ����
�F(xi��n)�ڱ���ˮ�a(ch��n)�B(y��ng)ֳ�I(y��)���ü��s����ʽ��(j��ng)�I(y��ng)�����s���B(y��ng)ֳϵ�y(t��ng)�����~�r���B(y��ng)��(sh��)����,�ܶȺܴ�,��Ҫ���]�ɂ�(g��)��(w��n)�}:����,�~�rֻ�ܵõ�����������Ȼˮ��ʳ��,��˱�횏���Ʒ����еõ������еĠI(y��ng)�B(y��ng),���������Ȼ��ЧɫԴ���ҳɷֲ���(w��n)��,��(d��o)���~�r���ܫ@�ó������}����,���wɫ�_(d��)���������Ҫ��; �ڶ�,�~�r���ڸ��ܶ�Ⱥ��,����������(y��ng)��,���Ү�(d��ng)��Ⱦ���l(f��)���r(sh��)���������ٶȸ��졣
A.�ɸ���ˮ�a(ch��n)��(d��ng)���Ƥ�w�ͼ�����ɫ������^�p�~��^�p�ԣ�
B. �����ˮ�a(ch��n)Ʒ����Ʒ�r(ji��)ֵ���M�����M(f��i)�ߵ����M(f��i)������
C.���Ԍ�(du��)ˮ�a(ch��n)��(d��ng)���M(j��n)��ҕ�X�̼������M(j��n)�zʳ��
�~�ɫ�ؼ�(x��)��
�~��wɫ��N���� ,ɫ����N�������@Щ������Ҫ�ɟo(w��)��(sh��)Ƥ�wɫ�ؼ�(x��)���a(ch��n)���ġ��F(xi��n)��֪���~�ɫ�ؼ�(x��)���ɷֳ����ЎN :
1����ɫ�ؼ�(x��)�� ,���к�ɫ�w����
2���Sɫ�ؼ�(x��)���c�tɫ�ؼ�(x��)�� ,��ԭ���|(zh��)�к��мtɫ���Sɫ��(n��i)���
3���B�S�ؼ�(x��)���ֽк���ؼ�(x��)�����ɫ�ؼ�(x��)�����ʼ�(x��)����
�����~�����ɫ���������õĻ�����w�{����(l��i)�s���� ,����}���������� (����ɫ�� ) ,�Sɫ��� ,��������Լ����ʺ����ʼ��F(tu��n)��
��ˮ�a(ch��n)�B(y��ng)ֳ�a(ch��n)���@��������ͬ�r(sh��)�����M(f��i)�ߌ�(du��)��(d��ng)����wɫҲ�_ʼ�v������(l��i)�����Ը����B(y��ng)ֳˮ�a(ch��n)��(d��ng)���wɫ�Ć�(w��n)�}�ѳɞ�Ŀǰ�������(gu��)ˮ�a(ch��n)��(d��ng)��I(y��ng)�B(y��ng)�W(xu��)�ҵ�һ��(g��)��Ҫ�о��n�}�������(l��i)���о�������Ӱ�ˮ�a(ch��n)Ʒ�wɫ��������ˮ�ء���ϡ���B(y��ng)�ܶȡ��~�w�Nʹ�С�ȡ����������֬����֬���ԾS���غ�ɫ�ص��о��^�࣬ɫ��������}���ط�����о���ࡣ
ˮ�a(ch��n)��(d��ng)����ɫ���h(hu��n)���c�wɫ�����h(hu��n)���l����(hu��)Ӱ��~��ɫ���γ� ,�~Ҳ��(hu��)��׃�wɫ�m��(y��ng)������ı�������(j��)Ellis��1997���(b��o)�� ,���l��wɫ׃���֞�ɫ����ɫ�ؼ�(x��)����(n��i)�Ƅ�(d��ng)���γɵĿ��ّ�(y��ng)��׃���ͼ�(x��)���gɫ�����|(zh��)������ľ���׃�����Gartner(1986)�w�{�ˮ�(d��ng)�r(sh��)��Ȼ����F(xi��n)���wɫ���������l��Y�� ,�l(f��)�F(xi��n)�wɫ�����Ă�(g��)�w��Ҋ���ذ��\ˮ̎ ,�J(r��n)����� (������ )�c�wɫ�������P(gu��n)�����(l��i)�C��(sh��)���⾀��ݗ���ܴ̼����Һ�ɫ�ص����ɡ�
�֓�(j��)Denson(1997)��(b��o)�� ,�������37d���ӹ��Տ�(qi��ng)�� ,��ʹ�����²��ְ���Į������ �wɫ���� ,��׃�B(t��i)�r(sh��)�����^(gu��)��(qi��ng)Ҳ��(hu��)��(d��o)�°���
ˮ�a(ch��n)��(d��ng)����ɫ��
�И���(j��ng)�c�wɫ����
Ӳ���~�ɫ���γ�����(j��ng)�̓�(n��i)���ڵ��{(di��o)�ء�Fujii��Oshima(1986) �о��l(f��)�F(xi��n),ጷŃ���Ӱ�����(j��ng)ԭ�����X���w���ڴٺ��ؼ�(x��)�����ɼ���,�M(j��n)��Ӱ��wɫ���I�����ء����I�����غͶ�Ͱ��ȃ���Ӱ��Ǒ�(y��ng)���r(sh��)�{(di��o)�������wɫ׃������Ҫ���ء�����Ӱ�-��-���D(zhu��n)��ø�c�wɫ�ľ���׃�����P(gu��n) ,���Ұ���ø�t�Ǻ�ɫ�����ɵ��P(gu��n)�Iø����ɫ���ں�ɫ�ؼ�(x��)����(n��i)�ĺϳɷ֣��� :�Ұ����������ɶ��u�������� (DOPA) ;�������ɶ�Ϳ��� ;��Ϳ������Ұ���ø�������¾ۺϳɺ�ɫ�ء�ͨ�� ,�~�ı��������Ұ���ø�ĺ����ͻ����^����� ,�wɫҲ�^���Щ�wɫ�����~��Ұ���ø�Ļ���Ҳ�ܸ� ,�����ؼ�(x��)���Л]�к�ɫ�� ,�����Ǵ�
�ڶ�N��ʽ���Ұ���ø,�����е�̎�����Ơ�B(t��i)��
�I(y��ng)�B(y��ng)�c�wɫ����
�˹���B(y��ng)�İ����l���L(zh��ng)�ʹ������ ,������ɰ������|(zh��)���cɫ���γ����P(gu��n)�ĠI(y��ng)�ɷ֡�Yamamoto��(1992)��(b��o)�� ,�zʳ�����Ѻ���Ȼ���΄�(d��ng)������Ұ�����͡�Estevez
(1996)��Miki��(1990)���D�� (݆�x���u�x )�������m����n-��ϵ�и߶Ȳ��֬���� (HUFA) ,�����Ƕ�ʮ��̼��ϩ�� (DHA)����ʮ̼��ϩ�� (EPA)�;S���أ� ,ʹDHA��
EPA��ֵ��1.36 ,Ҳ����Ч�A(y��)�������Һ����ҵİ�����
������(sh��)�(y��n)���� ,�S���أ��ͣ� ,DHA ,EPA ,��ʮ̼��ϩ��(ARA)����֬���Ƿ�ֹ��Ŀ�~�wɫ���������|(zh��)���A(ch��)������ ,�@Щ���|(zh��)�Ĵ_�Д�(sh��)�����m�˱��� ,����(sh��)�(y��n)�l���IJ�ͬ ,Ŀǰ�Пo(w��)һ�µĽY(ji��)Փ��
�A(y��)����Ŀ�~�wɫ��������Ҫ;���O��1998�����ĠI(y��ng)�B(y��ng)�W(xu��) (��ϵ�EPA�c�S���ؽM�ɼ����D
�ď�(qi��ng)�����g(sh��)�� )������W(xu��) (��(x��)��ɫ�غ���(j��ng)��(n��i)���� )�����B(t��i)�W(xu��) (���B(y��ng)���B(y��ng)ֳ�h(hu��n)���� )�C�����wɫ������ԭ���A(y��)����ʩ��
݆�x���u�x���w�ͳ��w��С�m�� ,�m���ԺͿɵ��ԏ�(qi��ng) ,���ڏ�(qi��ng)�� ,�I(y��ng)�B(y��ng)�r(ji��)ֵ�� ,��Ŀǰ�~ͼך�������г��õĻ��D�� ,Ҳ�ǽ�(j��ng)�^(gu��)��(qi��ng)�����A(y��)����Ŀ�~�wɫ��������Ч;��֮һ������������h(hu��n)����ͬ ,��ͬ�^(q��)����Ȼ���D�ϠI(y��ng)�B(y��ng)�ɷֺ��A(y��)���wɫ������Ч��Ҳ������
Solbakken�Ȍ�(sh��)�(y��n)�C��(sh��),�����������0.1��10 -������ĵ�נ����� (T4)�ܴ��M(j��n)ӹ�l��׃�B(t��i) ,��ֹ�wɫ����������(j��)�~�wɫ���γɵęC(j��)�� ,��Ͷιȫ�r(ji��)��Ϻͼӏ�(qi��ng)��B(y��ng)������ͬ�r(sh��) ,��������m������ɫ���γɵ�������ø������Ұ���ø��T4,������һ�N���������B(t��i)��Y(ji��)���A(y��)�����l��wɫ����;��֮һ��
���l�wɫ�������H�c�С����~����и߶Ȳ��֬����ķN���������P(gu��n) ,߀�c�H�~�ĠI(y��ng)�B(y��ng)���P(gu��n)������ͬ�����a(ch��n)�l�������� ,��ͬ���ε��ѷ��������~�wɫ�����ʸ��� ,��ԭ������ڴˡ���ͬ��׃���������� ,�z������(du��)�������~�wɫ�����ʵ�Ӱ��@���ش��ڌ�(du��)���L(zh��ng)�ͳɻ��ʵ�Ӱ푡���� ,�A(y��)�����l����(y��ng)���H�~�I(y��ng)�B(y��ng)ץ�� ,���Ǽӏ�(qi��ng)�H�~�ı���֬����I(y��ng)�B(y��ng) ,�����x��Ұ���H�~��
1.ˮ�a(ch��n)��(d��ng)��ɫ�ؽM��
�Ļ��W(xu��)�Y(ji��)��(g��u)��� ,���¿ɷ֞飺��}����Ⱥ��đ֭ɫ��Ⱥ����-����ϵɫ��Ⱥ����ɫ�ء����ϵɫ��Ⱥ������ɫ�ء��������q�~���\�������~��з����(du��)�r�ȼך���wɫ����ɫ�ļtɫϵɫ���Լ��~�ı�Ƥɫ ,��Ҫ���ǁ�(l��i)����}����Ⱥ��ɫ�ء�
ˮ�a(ch��n)��(d��ng)����ɫ��
��ɫ���ā�(l��i)Դ
�ܶ���(d��ng)���ֲ�ﶼ���и��N��Ȼɫ�أ��x���~ļ��⡢Ƥ�w�����Sɫ����ɫ�ͼtɫ���@Щ�ɫ��(du��)�^�pˮ����(d��ng)������~���\�����Լ������^�߽�(j��ng)��(j��)�r(ji��)ֵ���q�V�~����(du��)�r��������Ҫ���~�S��(���u����-���}����)���r����(���u��-��ͪ����-���}����)�ͽ��S��(��ͪ����-���}����)��ʹ�q���~ļ��⡢Ƥ�w�������ּtɫ����ɫ��ֲ���������������ɫ�ز�����ʹ�q�~�@���������ɫ����Ҫ��ֲ����}�������~�S�غ������S�أ�������Ҫ��������ޣ���S��������С�����rз�������r���أ�����ɫ�������Á�(l��i)Դ��
��}����
Ŀǰ��֪�Y(ji��)��(g��u)����}������ 600��N ,�������� 8��(g��)����ϩ��λ�M�ɵ�һ�̼?x��)仯���P�����������?��ܗ�p�I�L(zh��ng)���ƻ�����)�������Y(ji��)��(g��u)�Ƿ��Ѽt�� ,������}�����������������仯��Ó�䡢�h(hu��n)���Լ�̼�����š�����������ġ������(sh��)�y����ˮ ,��������֬���ЙC(j��)�܄��� ,�������ᡢ���Ȼ��R���ȷ���Һ�@ʾ���{(l��n)ɫ��һ����}�����ǣ�40 ���� ,��Ҳ���ڸ���}���� (��45�ͣ�50 )�ͽ������}���� (���30 )��
��}���� (Carotenoids)�ǏV����������Ȼ���еĸ��N��(d��ng)ֲ�P�����w�е�һ�ɫ�ء����~���Ҫ��(ch��)������Ƥ�w���~�[������ȽM���С���}���ؿɷ֞��� :
һ���̼?x��)��?,ֻ�ɣá��ȽM�� ,�Q����}���� ;
��һ��������� ,�ɣá��ȡ��ϽM�� ,�Q���~�S�ء�
��-���}����(���S���أ�ԭ )�Ǻ��}���صĴ��� ;
�S�w�غ��r���t���~�S�ص���Ҫ������
��}���صķNͷֲ���ˮ�a(ch��n)��(d��ng)���г�Ҋ����}������ ��- ���}���ء��S�w�ء�
�����S�|(zh��)�����~�S�|(zh��)���r���صȡ��~ؐ��wɫ֮�tɫϵɫ����Ҫ���r���� ,?�~��Ƥ��ɫ����Ҫ�ɷ��ǽ��~�S�|(zh��)����Ȼ������Ƥ����}���طֲ� :�r����ռ 60 %��
���~�S�|(zh��)ռ 2 0 %���S�w��ռ 1 5%�������S�|(zh��)ռ 4%����- ���}���غͽǼt�طքeռ 2 %�� 4%���� ,߀������������}���ء��z�X���ı�Ƥ�� ,�r����ռ 80 %����
�~�S�|(zh��)ռ 1 5%�������S�|(zh��)ռ 2 %,�Ǽt�ء������S�|(zh��)���������S�|(zh��)��ռ 1 %���� ,�S�w��Ҳ�������ֲ���
�ڽ����ı�Ƥ��,�r����ռ 75%�����~�S�|(zh��)ռ 2 0 %���Ǽt��ռ 3 %���S�w��ռ 2 %���� ,߀������������}���ء����~���\���� ,�����r���ء������S�|(zh��) (���w��(n��i)
�ϳɞ��r���� )��ɫ��Դ�Ĵ��� ,��ʹ��ʬF(xi��n)�tɫ��(qi��ng)���w���ɫ ,�����S�w�����w��(n��i)�ĺϳ����ö�ʹ�w���ʳ�ɫ��
��Ȼ�q�~���ɫɫ�����r���؞��� ,߀���нǼt�ء��S�w�صȡ�з���׳��˺��мך��|(zh��)���o(w��)�C(j��)�}�͵����|(zh��)�� ,߀���ж˻���tͪĺ��}��ɫϩ�ɫ�� ,���r���غ� ��-
���}���صȡ�
��}���صā�(l��i)Դ
ͬ������(d��ng)��һ�� ,ˮ�a(ch��n)��(d��ng)���w��(n��i)���ܺϳ���}���� ,��������}����ֻ������еõ���Ŀǰ���ӵ�ˮ�a(ch��n)��(d��ng)������е���}������Ҫ�Ѓɴ��һ�����Ȼ��}���� ,��Ҫ��һЩ������}���غ��~�S�صȵ��S���t����ɫ�Ą�(d��ng)ֲ������� ;�����ס����}�������r����ĸ��������O����ȡ�� ����Ȼ��}��������Ҫ�֞���}������~�S��� ��
���}�����Ǻ��Ўׂ�(g��)�����ϩ��λ�Ļ����� (������ ) ,�ж�N����(g��u)�w�������Ԧ����¡��� 3�N�Ҋ ,�������� �� -���}��������Ҫ ,������ߡ��� -���}���ؾ��оS���أ����������ܡ��mһЩ�~� (�����t�c(di��n)�q )���܌� �� -���}�����D(zhu��n)����S���أ� ,����Щ�~�(���|�~�����_�_���~�� )�t�܌� �� -���}�����D(zhu��n)����S���أ���ͬ�r(sh��) ,һЩ��(d��ng)���w�е� �� -���}������ֱ�ӻ��g�ӵ��c��ͪ������ ,�ڄ�(d��ng)�����ֳ���(d��ng)����������������ܡ�
�~�S�t�Ǻ��}���صĺ��������� ,���Դ���ȩ��ͪ�������ʽ���� ,�����ڼ״����Ҵ���ʯ���ѡ��~�S�ض���(sh��)�������оS���أ��Ļ��� ,���ұ���(d��ng)���w���c�����պ� ,���ܱ���ԭ��(l��i)�ķ��ӽY(ji��)��(g��u)���ɫ ,��������Ƥ�w��֬���M�����г��e����Ҋ���~�S��������� 5���  1 )�~�S��(Lutein) ,�� 3 ,3��-���u�� -�� -���}���� ; (2 )�����S��Zeaxanthin) ,�� 3 ,3��-���u�� -�� -���}���� ;(3 )�[�S��(Cryptoxanthin) ,�� 3 -�u�� -�� -���}���� ; (4)�̽��S��(Citroxanthin) ,�� 5,8-�h(hu��n)�� -�� -���}���� ;(5)�r�S��(Astaxnthin) ,�ֽ��r���� ,�� 3 ,3�� -���u�� -4,4�� -��ͪ �C�º��}���ء������r�S�؞�ˮ�a(ch��n)��(d��ng)���B(y��ng)ֳ�еij���ɫ�ء� 1 )�~�S��(Lutein) ,�� 3 ,3��-���u�� -�� -���}���� ; (2 )�����S��Zeaxanthin) ,�� 3 ,3��-���u�� -�� -���}���� ;(3 )�[�S��(Cryptoxanthin) ,�� 3 -�u�� -�� -���}���� ; (4)�̽��S��(Citroxanthin) ,�� 5,8-�h(hu��n)�� -�� -���}���� ;(5)�r�S��(Astaxnthin) ,�ֽ��r���� ,�� 3 ,3�� -���u�� -4,4�� -��ͪ �C�º��}���ء������r�S�؞�ˮ�a(ch��n)��(d��ng)���B(y��ng)ֳ�еij���ɫ�ء�
ˮ�a(ch��n)��(d��ng)����ɫ��
�ڶ���ǻ��W(xu��)�ϳɵ���}���ء����W(xu��)�ϳɵ���}������������270�N ,ͨ����ˮ�a(ch��n)��(d��ng)���B(y��ng)ֳ�Б�(y��ng)�õĞ�һЩ�����ĺϳ���}���� ,�� �� -���� -8�� -���}����ȩ(BAC)����-���� -8�� -���}���������� (BACE)�������S(CHX)�����S�� (CTX)����ަ�S�ء����ͪ��.
ˮ�a(ch��n)��(d��ng)����ɫ��
ˮ�a(ch��n)��(d��ng)����wɫ������(j��ng)�̓�(n��i)����ϵ�y(t��ng)���Ƶ� ,ͬ�r(sh��)Ҫ�ܵ��h(hu��n)����Ӱ푡�ˮ�a(ch��n)��(d��ng)��ɫ�{(di��o)�ij��F(xi��n)�c��}���ء���ɫ�ء���ʺ��B���ʵ�������(f��)�s���P(gu��n)ϵ��ˮ�a(ch��n)��(d��ng)��Ƥ�wɫ������Ҫ�Ǻ��}���ض������S�wɫ���r���ء������S�|(zh��)����}���ء���һ�����B(y��ng)ֳ�l���� ,һЩˮ�a(ch��n)��(d��ng)����wɫ�����|(zh��)�c��Ȼ�l���������ͬ�N��(g��)�w�����Щ׃��������ԭ�� ,����������������h(hu��n)���IJ�ͬ ,��Ҫ���B(y��ng)ֳ�����D����ȱ���~��x�ϳ�ɫ�������ǰ�w��ɵġ�
ˮ�a(ch��n)��(d��ng)����ɫ��
������r�� ,�~���rƤ�w ,�Լ�������ѵ�ɫ�ɴ�(l��i)������е���}���� ,�����Ձ�(l��i)����}������ԭ��(l��i)����ʽ���D(zhu��n)����������}�������~�w��(n��i)���e���@ʾ�����е��wɫ����ɫ��
ˮ�a(ch��n)��(d��ng)����ɫ��
2.�о�ˮ�a(ch��n)��(d��ng)��ɫ�ص�һ�㷽��
2. 1��ɫ����ȡ��һ���Ȍ���(d��ng)��M����{ (��{�r(sh��)����һ�����ğo(w��)ˮ�����c ) ,�ٲ����ЙC(j��)�܄� (��ͪ���ͪ��ʯ���ѻ��ͪ�ͼ״��� )�����ϳ�����������íh(hu��n)�����c��ƿ�M�ϳ���г����b�ó�������������� ,Ȼ���x�� ,��ɫ�ش���������Һ�С�
ˮ�a(ch��n)��(d��ng)����ɫ��
2.2��ɫ�ط��x�c������
һ����ü����������ӌ���������������ЧҺ��ɫ�V�͚���ɫ�V�ȷ������ͼ�������(l��i)�f(shu��) ,���ñ�ͪ��ȡɫ�� ,��ô��ȡҺ��(j��ng)��s���c(di��n)���ڞV���� ,Ȼ�������Ȼ�̼�ڲ���������չ�Ӽ��� ;�ͱ��ӌ�����(l��i)�f(shu��) ,��(y��ng)���ù��z���Ʊ��Ӱ� ,Ȼ���c(di��n)�� (�ý�(j��ng)��s��ɫ�ر�ͪ��ȡҺ��ɫ��ʯ������ȡҺ ) ,����չ�ӄ� (ʯ���ѡñ�ͪ�î����� 68��2 8��4��ʯ���ѡñ�ͪ�������� 90��1 0��0 45)չ�Ӽ��� ;�������tһ�����������������b�� ,��ʯ���ѻ����͡ñ��� 1
0��1�� 1��2�Ļ��Һ�M(j��n)��ϴÓ��Һ��͚���ɫ�V�tһ�������M(j��n)һ�����ԺͶ����y(c��)����
ˮ�a(ch��n)��(d��ng)����ɫ��
3.ɫ����ˮ�a(ch��n)��(d��ng)���w��(n��i)�Ĵ��x;��
ɫ����ˮ�a(ch��n)��(d��ng)���w��(n��i)�Ĵ��x;�� ,Ŀǰ߀��ʮ����� ,����Ҫ�����ڌ�(du��)��}���ص��о��� ,����ɫ��߀�����о���Hate��������������S�غϳ��r���صĴ��x;�����f(shu��) :�����S�ء���- ���}���������� 4 ���������S�ء��r���ء�߀�ЌW(xu��)�����M(j��n)һ���J(r��n)����}�������ձ���(du��)�r�w��(n��i)�ЃɷN���x;�� ,�� ��- ���}���ء����S�ء���đͪ�����S�ء��t�S�ء��r����;�����S�|(zh��)�� 4 ͪ�����S�|(zh��)���r���ء����Nɫ����ˮ�a(ch��n)��(d��ng)���w��(n��i)�Ĵ��x;���S��(d��ng)��NIJ�ͬ����ͬ ,���P(gu��n)ɫ����ˮ�a(ch��n)��(d��ng)���w��(n��i)�Ĵ��x;���ʹ��x;���е��{(di��o)���c(di��n)���{(di��o)������ ,߀�д����M(j��n)һ�������о���
ˮ�a(ch��n)��(d��ng)����ɫ��
3.ɫ����ˮ�a(ch��n)��(d��ng)���w��(n��i)�Ĵ��x;��
Goodwin���J(r��n)���r�������ձ���(du��)�r�w��(n��i)���x�ЃɷN:
һ;���� : �� -���}���ء��r���� ;
��һ;���t�� :�����S�|(zh��)���r���ء�
Liao wenliang��(b��o)�� ,��}�����е������S�|(zh��)��(du��)����(qi��ng)�r�wɫ������Ҫ����������?y��n)?�� -���}��������;���L(zh��ng) ,Ч�����������S�|(zh��)�ľ��ʡ�
4.ˮ�a(ch��n)��(d��ng)����ɫ���о��M(j��n)չ
��(du��)ˮ�a(ch��n)��(d��ng)����ɫ�����о� ,���ā�(l��i)�f(shu��)�LJ�(gu��)���������(gu��)��(n��i)��������(du��)��(j��ng)��(j��)��(d��ng)���о���(b��o)���� ,��(du��)�^�p��(d��ng)���������^�p�~�о���(b��o)���١��ߙn���^�p�~���Ǐć�(gu��)���M(j��n)�� ,���B(y��ng)ֳ֮���wɫ�O����ȥ��ʧȥ���^�p�r(ji��)ֵ��Ŀǰ ,��(gu��)��(n��i)��(du��)�M(j��n)���^�p�~����ɫ���wɫ����߀�ஔ(d��ng)���y ,�б�Ҫ��(du��)ˮ�a(ch��n)��(d��ng)����ɫ���M(j��n)�������о���
4.1��Ȼ��ɫ��
4.1.1����{(l��n)��;G�匦(du��)�~�r���������õ���ɫЧ�� ,��(du��)�r�w��(n��i)�r���ص���������Ч����֬����������ں��V�~�w����ɫ�س��e ,С��������}����ɫ�ص���Ч��(l��i)Դ ,��Ч�����ںϳ�ɫ�ء��þG�匦(du��)�Ѻ��V�M(j��n)��Ͷιԇ�(y��n) ,�о��G�匦(du��)�Ѻ��V����ɫ�ص�Ӱ� ,��(j��ng)�^(gu��)����Ͷι�� ,�M(j��n)������y(c��)ɫ ,�Y(ji��)������ ,�Ѻ��V����ɫ�ص������������ɫ���� ,��ɫ�ʺ����������p�͡��ú� 3 %����������ι�B(y��ng)�߹�(ji��)��(du��)�r ,�Y(ji��)���l(f��)�F(xi��n)��(du��)�r�^�ؼ��е���}���غ����@������ ,���sδ�y(c��)�������S�|(zh��) ,�������е���Ҫ��}���ر��߹�(ji��)��(du��)�rѸ�ٴ��x�D(zhu��n)�������r�S�|(zh��)��
��(j��)�˰l(f��)�F(xi��n)�C��(sh��) ,�B(y��ng)ֳ�߹�(ji��)��(du��)�r�ڲ��@����һ��(g��)��ǰͶι�������� 3 %���D������ߌ�(du��)�rɫ�غ�������Ч������
���ձ� ,���������������\���ͽ��~����ɫ�� ,�����L(zh��ng)�õĚvʷ����������ι�B(y��ng)�^�p�~ ,��Փ�Ǽtɫ�ص��~ (�����~�����~���t?�������~�� )߀�ǷǼtɫ�ص��~ ,�wɫ��(hu��)ͬ��׃���r�G���� ,�����L(zh��ng)��ֳ�������@���ӡ���(j��)�F(xi��n)���Y�Ϸ������� ,�����塢С�������������۶��Ǹ����~�r�wɫ�O�õ�ɫ��Դ ,��Ŀǰ��(y��ng)���^�����Ҫ�������塣�ڼt����������������� ,�������е������S�،�(du��)���~����ɫ������ ,���~�S��?z��)o���á�
�����~��������� 3 %- 6%�������� ,10W�����~�wɫ����ʮ�����@������������� 5%�� 10 %��������� ,��(du��)����ɳ���~Ͷι�S�� ,83d�� ,Ͷι���������ԇ�(y��n)�M ,��
Ƥ�wɫ�غ����@�����ڌ�(du��)�սM ,�����^���� ,�S�G���r�� ,�wɫ�����@���ơ�����ۡ��� -���}���غ��r������B(y��ng)�ձ���(du��)�r ,�Y(ji��)������ ,������ۺ������r���ص�Ч���ஔ(d��ng) ,�wɫ��ѡ��ݝ��iָ�� :�ձ����ϳ��r���~�r��ˮ����(d��ng)���D���������z�������ۼ��РI(y��ng)�B(y��ng)���з���ɢ���� ,�e���ڵV���|(zh��)�;S����ȱ����ˮ��Ч���Ȟ����@�������m��������~����a(ch��n)������������� ,�����~�w�wɫ ,���M(j��n)֬�|(zh��)���x ,��(ji��)�s�����|(zh��)��
�ܶ��W(xu��)���о��C��(sh��),�������������~�w��� ,����߹�(ji��)��(du��)�r���wɫ ,���M(j��n)��(du��)�r���L(zh��ng)������������������D��ι�B(y��ng)�\�� ,ʹ�\���wɫ�����r�G ,�����������L(zh��ng)��������������D��Ͷι���A�q��з (Eriocheir sinensis)з��,����ʿ����25.2 %,׃�B(t��i)���� 1-2��,����������������w�������D�� ,���C��������w��ֲ���Ե����|(zh��)�Ĕz�롣������߀�ܴ��M(j��n)��з�����L(zh��ng)�ͼ����з���wɫ;����ͬˮ�ؼ�Ͷ�D�� (Ͷι��ռ�w�صİٷֱ� )�ȗl���� ,�S������ɷ�Ͷι�������� ,��з�w������ ,��з�wɫ�����Ͷι�������� 6%���D�Ϟ�����m(�����,2001)��
�����~��������� 3 %- 6%�������� ,10W�����~�wɫ����ʮ�����@������������� 5%�� 10 %��������� ,��(du��)����ɳ���~Ͷι�S�� ,83d�� ,Ͷι���������ԇ�(y��n)�M ,��
Ƥ�wɫ�غ����@�����ڌ�(du��)�սM ,�����^���� ,�S�G���r�� ,�wɫ�����@���ơ�����ۡ��� -���}���غ��r������B(y��ng)�ձ���(du��)�r ,�Y(ji��)������ ,������ۺ������r���ص�Ч���ஔ(d��ng) ,�wɫ��ѡ��ݝ��iָ�� :�ձ����ϳ��r���~�r��ˮ����(d��ng)���D���������z�������ۼ��РI(y��ng)�B(y��ng)���з���ɢ���� ,�e���ڵV���|(zh��)�;S����ȱ����ˮ��Ч���Ȟ����@�������m��������~����a(ch��n)������������� ,�����~�w�wɫ ,���M(j��n)֬�|(zh��)���x ,��(ji��)�s�����|(zh��)��
�����庬�S���ĵ����|(zh��)������֬���ᡢ���ǡ��S���ء���Ȼɫ�ء��V���|(zh��) ,�Լ����L(zh��ng)���Ӻ̓�(n��i)Դ��ø��������Գɷ֡����������I(y��ng)�B(y��ng)�ɷֺ��������Գɷ��ڽM�ɱ������S������ķN��B(y��ng)ֳ�h(hu��n)���l���IJ�ͬ�����в���Д�(sh��)��(j��)���� ,�������еĵ����|(zh��)�������_(d��) 70 %���� ,�ஔ(d��ng)�ڴ� 1.7�� ,ţ��� 3.5�����䵰���|(zh��)�����������ֹ����ֲ����ʳƷ������О���� ,��������谱���Ậ���dz���ȫ��
�����忂֬�������� 6 %�� 9% ,֬���Ậ���� 55%���� ,���� 65%���֬���ᡣ���֬�������Ա���֬�����灆���ᡢ������ͻ�����ϩ�Ậ�����_(d��) 25%���ϡ��@Щ���DŽ�(d��ng)���w���x������Ҫ�����������|(zh��)���ʮ̼��ϩ�� (EPA)����ʮ��̼��ϩ�� (DHA)��ǰ������ (PGE)����ϳɵ�ǰ�w ,�@Щ֬����������ֲ��������к��������١��ھS���غ���Ԫ�ط��� ,�������оS���طN��ࡣ�����帻����-�� �}���ء������S�|(zh��)����đϩͪ����}���� ,����VB12�����İl(f��)�F(xi��n)�o�I(y��ng)�B(y��ng)�玧��(l��i)�@ϲ ,һ��ֲ��o(w��)��ͨ�^(gu��)��������γ�VB12,��VB12�ĺ����O�͡��������к����S�����˼���(d��ng)�����ĵV���|(zh��)Ԫ�ء����⣬������߀���и��������ǡ��\��Ԫ�ص�����������(j��)�@һ���c(di��n) ,���B(y��ng)ֳ��������^(gu��)���п�ͨ�^(gu��)���������\�͵�ĝ�� ,ʹ�������������\�͵�ĺ���������� ���Դ��_�l(f��)���������ߵ⡢���\�������塣�����帻��һЩС���Ӷ��ǡ������ǡ�����A���������������|(zh��)��
�������������Գɷ�����l(f��)�F(xi��n)����Ҫ�����L(zh��ng)���ӡ���������ǡ����{(l��n)�ء����w��߰�̓�(n��i)Դ��ø�ȡ������嵰���аl(f��)�F(xi��n)�ĔM���L(zh��ng)������һ�N�ɴ̼����w��(x��)�����L(zh��ng)�Ķ��� ,���д����L(zh��ng)���� �����{(l��n)������������һ�N��Ȼ�ɹ�ɫ�ص����|(zh��) ,�����ڸ��N�������ӵĘ�(bi��o)ӛ������ ,���{(l��n)��Ҳ����ɫ���YԴ�_�l(f��) ,�䌍(sh��)�{(l��n)�弚(x��)����ԭ���|(zh��)��������һ�N��Ȼ��ɫ�؎�(k��) ,�����~�G�ئ� ,�� -���}���ء��{(l��n)���S�غ��{(l��n)���~�S�صȡ�
4. 1. 2��ֲ�P����ȡ��
������}���ص�ֲ�� ,����}������ޣ���S���ס��Ϲϡ����?l��)ۡ���K�ݡ��f(w��n)�۾յ� ,߀���Sɫ�ͼtɫˮ����������ɫֲ�� ,����ʹˮ�a(ch��n)��(d��ng)����ɫ��Tunison�ȡ�Bss��Bawemfeind��Ellis��VRC��ӑՓ�����ɫ�ء��t���������������÷ۡ�з����r�����Լ��ϳɽ��S�،�(du��)�q�V�~��Ӱ푡��ռZ�� 5%����ޣ�ۿ��_(d��)��ˮ�a(ch��n)��(d��ng)�����ɫЧ������������������?l��)ۻ�?c��)���~�� 3 %�� 5%,ˮ�a(ch��n)��(d��ng)��Ҳ�ɫ@�����õ���ɫЧ������һ��������(n��i) ,�� -���}����������(qi��ng)���A�q��з�wɫ������ ,�����A�q��з�B(y��ng)ֳ��Ͷι�������D���ܴ�ʹ���A�q��з�ך���ɫ ,����������ǰ 2��(g��)��Ͷι�������D��Ч�����ѡ�
����ɫ��:�t������ÿǧ�˺���}���� 275��1650mg ,�������S���S�� ,����(gu��)������˾���a(ch��n)�ļt�G�G(KemGLO)���Ǐ�����ȡ�ļtɫ��}���� ,�����c���W�W��ϳɳȼtɫ����� ,�������q�V�~���r����ɫ���о��l(f��)�F(xi��n)���~�������t�����c����ɫ��Ͷι�����չ������������� ,�ְl(f��)�F(xi��n)���~�w���յ������t�ֽ�(j��ng)�������x�D(zhu��n)���� 4 ͪ�����t���Էe�� ,�f(shu��)������ɫ�،�(du��)���~�@ɫ��Ч.
���ɫ�� :
Canthaxanthin��(du��)�q�~���V�~�����~�����~�������|֮��ɫ����ʹ�� ,ÿ��������� 50����B(y��ng)���VЧ���@�������~���\�����wɫ�����������S�|(zh��)�^�ѡ����~ (Yellow Tail)��
Ethyl��APo8�� Carotenoate�^�� , Canthaxanthin��astaxanthin�������q�~���w�������������ɫ ,���߸������ڽ��~���\����Ϟ�w���ļtɫɫԴ ,�~�S�t�������~�w�����SɫɫԴ.
��}���� :�f(w��n)�۾�ÿǧ�˻���ۺ���}����2.35mg ,һ����������ϵ� 0.3 %,����(gu��)������˾���a(ch��n)�Ľ��W�W���Ǐ�����ȡ���Sɫ��}���ء����ձ� ,�������B(y��ng)ֳ��(ch��ng)���ý�K�� (Adoni sacuvaiis)������ȡ�Ľ��Sɫ��}��������ɫ�� (�����û���ֱ������ ) ,�� 1 %���µı������ӵ������ι�B(y��ng)���V ,һ��(g��)������ ,���V�ı�Ƥ�[�ׄt׃���Sɫ ,�ɫ�O������ ,�ஔ(d��ng)�ÿ����˷����H��(hu��)�������V�����|(zh��) ,���ҕ�(hu��)������I(y��ng)�B(y��ng)�ɷ� ,ʹ���V���|(zh��)���r�ۿɿڡ�
��}�����ڻ������~����е����m��������(y��ng)��400mg/kg���������~����������յ�ɫ�س��ٲ�������I(y��ng)�B(y��ng)���|(zh��)�D(zhu��n)����S���أ��� ,���ಿ�־����e���~�w���w�������������
��̎�����Ӽ�������}���غ��� ,ʹ���ɫ���ͬ�~�һ�� ,�ڻ����������L(zh��ng)�^(gu��)���� ,��}����Ҳ�ܴ�ʹ���ٰl(f��)�����ڴ��~���ѳ��г��e ,���M(j��n)���S�ij��� ,�������S�ɫ�����Գ�������~�w�����e�γɻ���ɫ ,�Ķ��������^�p�wɫ.
4. 1. 3����(d��ng)��
4. 1. 3. 1���͵Ȅ�(d��ng)�P��U���Ѫ�t�x��ˮ���tɫˮ����[�r����ʹ�~���wɫ�����G���������r�D����B(y��ng)�����r(sh��) ,�r���ռ�ı������w�� 10 %-40 %,һ����~�w�� 400g���ҵĴ����_ʼʹ�� ,��ɫЧ�����@���ú��б������r���w���ռZ�ι�^�p�~ ,�S�ռZ��}���غ����������~�wƤ�w�ļtɫ�ɱ����ļ���r��з��ĵϠ�����x�@Щ��(d��ng)��ך��к����r�S�� ,�������r��з���~��ˮ�a(ch��n)��(d��ng)�����ɫ�����r�����r������v����ι�B(y��ng)���~ ,Ҳ��������ɫЧ�� ,�����nj�(du��)�tɫ���Sɫ����ɫ�Ľ��~ ,��ɫЧ�����ѡ�ˮ��Ҳ����ʹ�tɫ���~���wɫ�����G�����҇�(gu��)���ÏU�����r�^���r���ӹ����r���� ,���~�r��������� 1 %��3 %��(du��)�����~�r�wɫҲ��һ��������.
4. 1. 3. 2���͵Ȅ�(d��ng)����ȡ�
�r���غ��r�t���Ǐĵ͵Ȅ�(d��ng)������ȡ����(l��i)����Ȼɫ�ء���ˮ�a(ch��n)��(d��ng)����������� ,���Ը�׃ˮ�a(ch��n)��(d��ng)����wɫ��
�r����(astaxanthin) �����r�S�|(zh��) ,��һ�N 3 ,3��- ���u��- 4,4��- ��ͪ��- �� ,�¡�- ���}���� ,��һ�Nͪʽ��}����,�����ʽ��C40 H52 O4,�����|(zh��)���� 596. 86,��һ�N��ϩ�ͻ����� ,�ʷۼtɫ ,�� 600��N��}�����е�һ�N������֬����,������ˮ,�������ȷ¡���ͪ�����Ͷ���̼���ЙC(j��)�܄����������� ,������׃?y��u)��r�t�� �����V���������������,�e��ˮ����(d��ng)����r��з���~���B���ë��,���@ɫ�����á������������ 40.70��100mg/kg���r���� ,���V�����[������~�ʹ������q��ɫ�س��e�ٶȼ���λ�S�~�NIJ�ͬ����ͬ ,���wɫ�ļ����ɫ�غ��������Ӆs����ӹ���ɵġ�
��������ǰ 20d�� ,Ͷι������r���صĻ���� (������ÿǧ���~�w��ÿ�� 10-100mg����� 0.3mg�r���� )��(hu��)ʹ�˹��B(y��ng)ֳ�����wɫ�cҰ��������ͬ���r�G��ɫ�ʡ��ú� 36.9��35.4mgȫ-��-�r����/kg��ȫ-�źͣ�-�r���ػ�� (�քe�� 64%��35%)�ăɷN�w�����Ͷι��B(y��ng)�ڵ�ˮ�еĺ��V(�w�� 0.4kg) ,��(j��ng)�^(gu��) 31d ,�~�����r���صĝ�ȏ������ 1.7Mg/kg��ߵ�6-7mg/kg��69d�� ,Ͷιȫ-��-�r���ص��~������}���غ�������Ͷι�r���خ���(g��u)�w������һ�M (�քe�� 10.0�� 8.6mg/kg)����(sh��)�(y��n)�Y(ji��)���r(sh��) ,Ͷιȫ-��-�r����һ�M���wɫ���@���^�t ,���^���������@�^�� (79%��(du��)64%)���@������- �r���خ���(g��u)�w����(y��ng)�����~����ɫ�ıȲ���ȫ-��- �r����.
���ձ���(du��)�r��������� 0.1 %���r���،�(du��)���ƌ�(du��)�r�wɫЧ����ѡ����ҳɻ����cɫ�؝�ȳ������P(gu��n)���ڃɷN�ض���(8�桢12�� )�oƽ�� 150���صı��O�t�c(di��n)�q Ͷι��0-192mg
�r�S��/kg����ϵ� 6�N��ϡ����L(zh��ng)��ƽ���w��320g(12��r(sh��)�� 102�� ,8��r(sh��)��126��)���ך� ,�u(p��ng)�r(ji��)�~��ɫ�ص��γɡ��Y(ji��)���r�S���c�~��ļtɫ�ȳ������P(gu��n) ,��(d��ng)�D�����r�S�صĺ����� 70mg/kg�r(sh��) ,�~�wɫ�س��e���S�r(sh��)�g���ƶ��ʬF(xi��n)ƽ��(w��n)��B(t��i)���c�i��������λ��� ,�~β����ɫ���γɸ����ܼ� ,���N�ض����~���ɫ�c�ض����L(zh��ng)�ʳ������P(gu��n)�� 8���12��l�������L(zh��ng)���q��ɫ���γ����@��
�����r�������G���ɫ ,�����c����(d��ng)�����خ��Y(ji��)�� ,�������ˮ�a(ch��n)����� ,���Ը����B(y��ng)ֳ�~�Ƥ�w�ͼ���ɫ�� ,�����~Ŀ������������� ,�r���،�(du��)�~����L(zh��ng)��ֳ���к���Ҫ������ ,�����鼤�ش��M(j��n)�~���ܾ� ,�p����̥�l(f��)���������� ,���M(j��n)��(g��)�w�����L(zh��ng) ,���ӳ����ٶȺ���ֳ�� ��
���V��(du��)�r�S�ص���ɫ�Ƚ��S�ؿ� ,���λϵ��(sh��)�� 1. 3�� ,Ͷι 100mg/kg���r�S�ػ���S�ؽo���V ,�Y(ji��)��Ͷι�r�S�صĺ��V��������}���صĝ���^�� ,�̼����~����wɫ�r(sh��)�r�S�����ӄ����� 36-37mg/kg����(sh��)�(y��n)�Y(ji��)�� 4�ܺ�,���~�wɫ���ַ�(w��n)������� ,�ڟo(w��)���ˮ�w����B(y��ng)���~,����������r�S�|(zh��)�ǫ@���r�G�wɫ���m�˷�����Ͷι�����r�S�|(zh��)��ϵĽ��~,��ɻ����@�����ڲ������r�S�|(zh��)��ϵĽ��~ ,����������r�S�|(zh��)��(du��)�~�w��?z��)o�@��Ӱ�.
�ڰ߹�(ji��)��(du��)�r��������� 50mg/kg�r�S�|(zh��)(��ʲ��˾ ) ,��(du��)�r����}��������� 318%,70%-90%����}�������r�S�|(zh��)����ʽ���ڡ��r�S�|(zh��)�ij����ڼך��и��ڼ����� ,�ɂ�(g��)���ں� ,���e���ڼ������_(d��)����(w��n)����ͬ�r(sh��)�ڼך��еij��eҲ�ڷ�(w��n)�����ӡ��о�����,�r�S��������е��m����������(y��ng)�� 20-40 mg/kg,��9-11�ܕr(sh��)�g�M(j��n)��Ͷι�wɫ��׃�����@�ÝM��Ч��.�r�t��������ϩ���͵��r�����c�����|(zh��)�ĽY(ji��)������ΑB(t��i)�V�������ڼך����(n��i)������ ,����һ�N����ΑB(t��i)Ҳ������ijЩ�~Ƥ�С��r�t�����r�����c�����|(zh��)���x��(j��ng)�������ɵġ��r�t��������˂����V���о��ļtɫϵ����ɫ�������r�t��ι�B(y��ng)���V ,���Եõ��tɫ�V�~�����~�S��ι�B(y��ng)���V ,�t���Եõ��Sɫ�V�~��ͬ�� ,�rз���������(y��ng)ʹ�ütɫɫԴ.
4.1.4�����
�����еĹ�ϼ�(x��)�� ,�����ĸ�ͼt�����ĸ��Ҳ��ˮ����(d��ng)��ܺõ���Ȼ��ɫ�������t�����ĸ���ӵ��q�~���\�~������� ,�Y(ji��)���r��������Ƥ�w�ͼ����д����e�� ,��ʹ�q�V
�~�wɫ�ʬF(xi��n)�tɫ���ù�ϼ�(x��)����B(y��ng)���~ ,����ʹ�tɫ�صij��F(xi��n)�ӿ� ,���~���wɫ���r�G���о����� ,�����ĸ���@������_�����r�������ʺʹ���� ,��������� 14.48%,�����
��� 21.66%,���ܸ����_�����r���wɫ ,�r������}������� 40.40 %,ʹ�俴��ȥ�����������РI(y��ng)�B(y��ng).
4. 2����Ʒ��ɫ��
4. 2. 1������Ȼɫ�����្����ɫ����Ŀǰ ,����Ȼɫ��Դ����ȡ����ɫ����ˮ�a(ch��n)��(d��ng)����ɫ�аl(f��)�]��������á���ʿ�_�Ϲ�˾���a(ch��n)�ļ����ؼt���������S ,����(gu��)������˾�Ľ��W�W���t�G�G ,��(gu��)��˹��˾��¶���� ,�����������ӄ��S���a(ch��n)���w�S�صȮa(ch��n)Ʒ���Ǐ���Ȼɫ��Դ����ȡ������ɫ�� ,�������������õ���ɫЧ����������ĸ�����a(ch��n)ʮ�N��}���� , ��Ҫ���r���غ� ��- ���}���� ,��Ұ�������r���صĺ�����ռ 40%-95%��Ŀǰ��(gu��)��W(xu��)���ڱ������������з��x��һ�N��t�����ĸ ,�c�t�����ĸ��� ,���r���خa(ch��n)��Ҫ�� 80�� ,���ҠI(y��ng)�B(y��ng)Ҫ���^�� ,���a(ch��n)�ٶ��^�� ,�Ķ�ʹ�r���ص����a(ch��n)�����_(d��)����Ʒ�� ,��������ˮ�a(ch��n)��(d��ng)��֮��ɫ
4. 2. 2�����W(xu��)�ϳɷ����a(ch��n)����ɫ�������W(xu��)�ϳɷ����a(ch��n)����ɫ���к� 40��(g��)̼ԭ�ӵĺ��}���غͷ��Ѽt�� ,30��(g��)̼ԭ�����ҵ� �� ���� 8�� ���}��ȩ (C33 H40 O)�� �� ����8 ���}������֬ (C32 H44O)�������S�|(zh��) (C33H44O) ,��ʿ�_�Ϲ�˾�ļ����طۼt (���r���� 5%��10%)�ȡ��������ӵ��~����� ,���и����wɫ������.
4. 2. 3���������ɫ�������Gͪ��һ�N�˹��ϳɵ����Լ��� ,��һ�������¿����ƴ��wǰ�~�����ټ��ط��� ,���M(j��n)�����|(zh��)�ϳɴ��x ,���������ܺϳ� ��-���}���� ,Ҳ���ܴ�ʹ�������|(zh��)�ϳ� ��-���}���� ,������Gͪ����ֱ�������~�w����}���غ�����������ʹ̎��һ���l(f��)���A�ε��~���F(xi��n)�����D(zhu��n) ,�ɴ��~׃�����~���������Լ��ؿ�ʹ�^�p�~���wɫ���r�G,�䌍(sh��)�|(zh��)�����Ի��γɻ���ɫ�ľ��ʡ�
5.Ӱ�ˮ�a(ch��n)��(d��ng)����ɫЧ�������� ��
��ͬ��ȡ���ͬ�N���ɫ����(du��)���Nˮ�a(ch��n)��(d��ng)�����ɫЧ����(hu��)�����c�ڌ�(du��)�����ճ̶ȼ������ʲ�ͬҲ������ͬ���B(y��ng)ֳ��Ʒ�N�����L(zh��ng)�h(hu��n)��������еĠI(y��ng)�B(y��ng)�ɷֺ͠I(y��ng)�B(y��ng)���ӵȵIJ�ͬҲ��(hu��)ʹ��ɫЧ�����^��������B(y��ng)ֳ�^(gu��)���и��N��ɫ����ˮ�a(ch��n)��(d��ng)��Ƥ�w����������еij��eЧ�����cɫ�ر������ԺͽM�����P(gu��n)�⣬߀�c�B(y��ng)ֳƷ�N���佡����r��ֳ�^(gu��)���������D�ϵĽM�ֵȶ�N�������P(gu��n)��
5.1 �D�ϽM�ɡ��ӹ��c�A��
a.�D���е���֬�������������S����E�����|(zh��)�ܱ��o(h��)ɫ�������Ɖģ���������ˮ�a(ch��n)��(d��ng)�(du��)ɫ�ص�����;
b.�D�����纬�^�ߝ�ȵ��}�;S����A�t��(hu��)���͌�(du��)ɫ�ص����գ�Ӱ�����eЧ����
c.�D���е����|(zh��)����͡�֬����������B(t��i)����}���غ������D���ІΌ��ȸ��N��?f��n)I�B(y��ng)���ӡ����Nɫ�ص������Ԡ�w����С����(w��n)���Լ��N�
d.��ϼӹ��r(sh��)�����ضȸߵ͡��r(sh��)�g�L(zh��ng)�̣�����A��l�����r(sh��)�g���Ƿ��Ⱦù���Ⱦ���(du��)��ɫ��Ӱ�.
e.���ԭ�ϵ���}���غ���ֱ��Ӱ푵���ɫЧ����ԭ�Ϻ���}���ض� ,�t��ɫЧ���ͺ�.
f. ��ϵă�(ch��)�棬����σ�(ch��)��r(sh��)�g�^�L(zh��ng)���t��ɫЧ��Ҫ��Щ�������@Щ�Ⱦ��cɫ����ˮ�a(ch��n)��(d��ng)���w���������еij��e���P(gu��n).��
5.2 ˮ�a(ch��n)��(d��ng)��ı���
(1)��ͬ�N���ͬƷ�N����ͬƷ�N��ͬƷϵ�Ą�(d��ng)����eɫ���������z
���ϵIJ����(du��)��}���صij��eЧ�ʲ�ͬ��
(2)��(d��ng)���Ԅe��ͬ��(du��)��}���صij��e����Ҳ��ͬ,һ�����ԏ�(qi��ng)�ڴ��ԣ�
(3)��(d��ng)���w��ͬ��λ���eЧ�ʲ�ͬ����ɫ�س��e�İнM�����x���ԣ�
(4)��(d��ng)���������B(t��i)����נ��ٷ���������ø�������������Ȍ�(du��)ɫ�س��e����(hu��)�a(ch��n)��Ӱ푡�
(5) ������B(t��i)��(du��)ɫ�صij��e��Ӱ푡���Ӱ푄�(d��ng)���������յļ����Ⱦ���(du��)ɫ�صij��e�����в�����Ӱ푡��B(y��ng)ֳ�^(gu��)����ˮ�a(ch��n)��(d��ng)���һЩ���������������������������������w��(n��i)�ļ����x���Ⱦ���(hu��)�����~�r��(du��)�D�ϵĔzʳ�ʣ�Ӱ�?zh��n)D���и��Nɫ�ص�����������������r���Ķ�Ӱ�ˮ�a(ch��n)��(d��ng)���w��(n��i)ɫ�صij��eЧ����
5.3 �B(y��ng)ֳ�l��
�B(y��ng)ֳ�h(hu��n)����ˮ�ء�������PHֵ���������������B(y��ng)ֳ�^(gu��)���е�Ͷι���m(x��)�r(sh��)�gҲ��(hu��)Ӱ�ɫ�صij��e�ʣ�Ӱ�ɫ�����w���������еij��e����Ӱ��~�w�����|(zh��)��ɫ�ɡ�
5.4 ��ɫ������������
��Ҫ��ָɫ�ر����Ę�(g��u)�͡������w����(g��u)�w����Ȼɫ��һ��������ԣ����ڄ�(d��ng)���w��(n��i)���e�ʸ��_(d��)100�������˹��ϳɵ�ɫ�ض��o(w��)�����ԣ�����e��Ҳ����(y��ng)�^�͡��ή���(g��u)�w�з�ʽ�^�ʽ��(w��n)�����ܸ���Ч����ɫ������ɫ��������Ҳ��Ӱ������������ʵ���Ҫ����֮һ��
����Ӱ�����ʹ����ɫ�����Ӻ����ɫЧ���y�Թ�Ӌ(j��)���@��Ҫ���B(y��ng)ֳ�߲������������ˮƽ���ӏ�(qi��ng)��(du��)�B(y��ng)ֳ��(ch��ng)�ĿƌW(xu��)������������a(ch��n)�ߑ�(y��ng)��(du��)ÿ�Nԭ�ϵ��~�S�غ����������������M(j��n)�Йz�y(c��)����(du��)�˹��ϳ���ɫ����(y��ng)�М�(zh��n)�_�Ĕ�(sh��)��(j��)�Y�ϲ��Ì�(sh��)�H�z�y(c��)��߀��(y��ng)�{(di��o)������Ј�(ch��ng)��(du��)ˮ�a(ch��n)Ʒ��ɫ�̶ȡ���λ���ɫ��Ҫ��ͬ������(du��)�Y��ˎ���_(d��)���������ɫĿ�ģ�������F(xi��n)��������������r�t��ι���V�ütɫ�V�~�����~�S��ι���V�t�á��S���V�ˡ�
6. չ������(du��)ˮ�a(ch��n)��(d��ng)����ɫ�a(ch��n)��Ӱ푵������Ƕ��� :һ��ȡ�Q��ˮ�a(ch��n)��(d��ng)�ﱾ�����z������ ;����ȡ�Q��ˮ�a(ch��n)��(d��ng)����̎�ĭh(hu��n)�� ;����ȡ�Q��ι�B(y��ng)ˮ�a(ch��n)��(d��ng)�����õ��D�� ;����ȡ�Q��
�˞�ע���ɫ�ػ��صȡ���(du��)ˮ�a(ch��n)��(d��ng)����ɫ���о� ,��Ҫ��ͨ�^(gu��)Ͷι��ɫ�ػ��ص��D�ϻ�ͨ�^(gu��)�h(hu��n)�����ӵĸ�׃��(l��i)Ӱ�ˮ�a(ch��n)��(d��ng)����wɫ����ɫ��Ȼ�������÷ֹ��ȷ��y(c��)ˮ�a(ch��n)��(d��ng)��ɫ����ȡҺ��ODֵ ,�Դ_���䌦(du��)ˮ�a(ch��n)��(d��ng)����ɫ��Ӱ푡�Ŀǰ ,��(du��)ˮ�a(ch��n)��(d��ng)����ɫ�C(j��)�����о�߀���w�\ ,�е�����߀̎�ڼ��f(shu��)�A�� ,�]����������ɫ�����w��(n��i)�Ĵ��x;�� ,Ҳ�]����������ɫ����ˮ�a(ch��n)��(d��ng)���w��(n��i)���x���D(zhu��n)���ͳ��e�@һϵ���^(gu��)���е��{(di��o)���c(di��n)���{(di��o)�����ӡ����� ,δ��(l��i)��(du��)ˮ�a(ch��n)��(d��ng)����ɫ���о���(y��ng)�����ڴ��x;�����{(di��o)������ ,ֻ���@��ˮ�a(ch��n)��(d��ng)����ɫ���о��ŕ�(hu��)��һ��(g��)���_(t��i)�A��
���⣬���ڌ�(sh��)�H��(y��ng)�õ���Ȼɫ��Դ�٣��ҳɷֲ���(w��n)�����o(w��)�����Cһ��Ч������ˇ�(gu��)��(n��i)���������ڻ��W(xu��)�ϳ�������}���غ͏���Ȼ���аl(f��)���۷e��ȡ�����^����ɫ�أ������Է�(w��n)����̎�����_�l(f��)���Ա��C��ɫЧ����ǰ�����-����-��-���}��ȩ��BAC������-����-��-���}��������BACF���������صȣ���������ʿ�_�Ϲ�˾���a(ch��n)�ļ����ؼt���������S������(gu��)������˾�Ľ��W�W���t�G�G����(gu��)��˹��˾��¶������
�S���،�(du��)ˮ�a(ch��n)��(d��ng)�。��������
1. �I(y��ng)�B(y��ng)���ߌW(xu��)
�S���I(y��ng)�B(y��ng)�W(xu��)�����ߌW(xu��)���t(y��)�W(xu��)�ȌW(xu��)�ƵĿƼ��l(f��)չ���˂�?c��)��?l��i)Խ�J(r��n)�R(sh��)���C(j��)�w�I(y��ng)�B(y��ng)��r�c���ߙC(j��)��֮�g�������е��P(gu��n)ϵ���@��ʹһ�T�µķ�֧�W(xu��)�ƵĮa(ch��n)�������I(y��ng)�B(y��ng)���ߌW(xu��)������һ�T������Ҫ��Փ�r(ji��)ֵ���R�����x�����d�W(xu��)�ƣ��ǠI(y��ng)�B(y��ng)�W(xu��)�����ߌW(xu��)�����߅���W(xu��)�ƣ����о���(du��)��������ϵ�y(t��ng),��ҪĿ�����о��I(y��ng)�B(y��ng)���|(zh��)��(du��)�C(j��)�w����ϵ�y(t��ng)�����L(zh��ng)���l(f��)���͌�(du��)���߹��ܵ�Ӱ푼���C(j��)��,�о��I(y��ng)�B(y��ng)���|(zh��)���D(zhu��n)�ơ����ա����úʹ��x����P(gu��n)ϵ,������(y��ng)�������g(sh��)��(du��)�I(y��ng)�M(j��n)���{(di��o)�صȆ�(w��n)�}.
���w��(l��i)�f(shu��)�����ߠI(y��ng)�B(y��ng)�W(xu��)���о�?j��)?n��i)����Ҫ�ǣ��о��I(y��ng)�B(y��ng)�c���ߵąf(xi��)ͬ���á��I(y��ng)�B(y��ng)�����c�������P(gu��n)ϵ���I(y��ng)�B(y��ng)�c����ϵ�y(t��ng)�ļ�(x��)���ֻ��ͼ�(x��)�����x�a(ch��n)����P(gu��n)ϵ�Լ������@Щ�����M(j��n)�РI(y��ng)�B(y��ng)�{(di��o)�أ��о���(y��ng)����B(t��i)�£����ք�(d��ng)��������߹��ܵĠI(y��ng)�B(y��ng)��Ҫ���������(l��i)�����I(y��ng)�B(y��ng)(Micronutrients)�e�ǾS���غ���Ԫ���c����ϵ�y(t��ng)�P(gu��n)ϵ���о��ڠI(y��ng)�B(y��ng)�W(xu��)�����ߌW(xu��)�̈́�(d��ng)���t(y��)�W(xu��)�ȌW(xu��)���ѳɞ�ʮ�ֻ��S���I(l��ng)��
�I(y��ng)�B(y��ng)���ߌW(xu��)��һ�T���d�W(xu��)��,�ڄ�(d��ng)��I(y��ng)�B(y��ng)�о��I(l��ng)��ȡ���˾�ɾ�.�����(l��i)���Q�������ߴ̼����@�N����������ӄ�,�����ɶ�N�������|(zh��)��(j��ng)��(d��)�صļӹ���ˇ�Ƶ�.���ߴ̼����ֿɷ֞����ߠI(y��ng)�B(y��ng)����ߴ��M(j��n)����ߴ̼������.���ߠI(y��ng)�B(y��ng)������ڻ������|(zh��)�м���������������Ķ�N�S���ء���Ԫ�ء�ijЩ���谱���ᡢ֬����.�����(l��i)���о�ʹ�˂��J(r��n)�R(sh��)���S���،�(du��)ˮ�a(ch��n)��(d��ng)�������ϵ�y(t��ng)������ҪӰ�,�e�ǾS����C���S����E���S����B6�ȡ�
2.ˮ�a(ch��n)��(d��ng)��I(y��ng)�B(y��ng)���ߌW(xu��)�о��ı���
�S��ˮ�a(ch��n)�B(y��ng)ֳҎ(gu��)ģ�����U(ku��)���s���̶Ȳ�����ߺ��B(y��ng)ֳ�h(hu��n)������څ��������(d��o)���B(y��ng)ֳ�~���rз���B(y��ng)ֳ�ڼ����l���l(f��)�������������^�ߣ��䲡����(w��n)�}Խ��(l��i)Խ�ܵ��˂����P(gu��n)ע����Ҫ����һ�l������W(xu��)�ί������~���rз�����ķ��������о�������~���rз�����Ŀ�������,��
�����~���rз����һ�l��Ч��;������(j��)Raa��(1992)���(b��o)���������ܽ���ˮ�a(ch��n)�B(y��ng)ֳ��(d��ng)�ﱩ�l(f��)������Σ�U(xi��n)�����F(xi��n)�������خ��ԏ�(qi��ng)��߀�]��һ�N�ܿ���N��������Ч���硣���⣬�����غ��������W(xu��)ˎ����Á�(l��i)����Ϳ����~���rз�ļ��Ը�Ⱦ����������ˎ������h(hu��n)����Ⱦ����(x��)����ˎ�ԵȆ�(w��n)�}��ʹ�䑪(y��ng)���ܵ����ơ���ˣ���(gu��)��(n��i)��һЩ�W(xu��)�ߏ����ߌW(xu��)�ǶȌ�(du��)�~���rз���������M(j��n)�����о���ԇ�Dͨ�^(gu��)��(y��ng)�����ߴ̼����̼��~���rз����������ϵ�y(t��ng)����ߙC(j��)�w���خ������߹��ܺ͌�(du��)�����ĵֿ������Ķ��_(d��)�����μ�����Ŀ�ģ��@���湤����ȡ����һ�����M(j��n)չ��
�~���rз�����ϵ�y(t��ng)��׃���ڝO�I(y��)���a(ch��n)�зdz���Ҫ����?y��n)�����ϵ�y(t��ng)�ēp����(hu��)��(d��o)���~���rз�������l(f��)��������(Maule et al.,1989)���I(y��ng)�B(y��ng)��r���J(r��n)���ǛQ���~���rз����������Ҫ����֮һ���I(y��ng)�B(y��ng)��(du��)�~���rз�Ľ��������������@����Ӱ푣����оS����(��VE��VC)��Ӱ��~���rз����ϵ�y(t��ng)����Ҫ�����ӡ��I(y��ng)�B(y��ng)����������Á�(l��i)��ֹ�������l(f��)�͜p�p��Ⱦ�ć�(y��n)����(Chandra,1996;Scrimshaw & SanGiovanni,1997)��
�I(y��ng)�B(y��ng)�،�(du��)ˮ�a(ch��n)��(d��ng)�����ߵ�Ӱ���Ҫ��Ӱ�����خ������ߡ�ˮ�a(ch��n)��(d��ng)��ķ��خ������ߙC(j��)�� ,����Ѫ���a(b��)�w���ܾ�ø���D(zhu��n)�F���ȷ��خ����wҺ���ӵĻ��(d��ng)�Լ����ɼ�(x��)���͆κ˼�(x��)�������ɻ��(d��ng)�� ,��(du��)��ˮ�a(ch��n)��(d��ng)��ֿ���ԭ�����u��Ⱦ������Ҫ���á���������(qi��ng)����������(qi��ng)ˮ�a(ch��n)��(d��ng)�(du��)��N��Ⱦ�Բ�ԭ�ķ��خ������߷���(y��ng) ,��ߙC(j��)�w��������
���w��(l��i)�f(shu��)���I(y��ng)�B(y��ng)�،�(du��)�~�����Ӱ푵��о���Ҫ�����ڷ��خ������ߺͼ�(x��)���錧(d��o)�������Լ����w�錧(d��o)�����ߣ�����(du��)�ڼך������Ӱ푵��о��t��Ҫ�����ڷ�����ø(PO)���ܾ�ø���������������ء���(x��)�����������ȷ��خ������ߵ��о���
�a(b��)��S���ص�Ŀ�ģ�
A.�M��ˮ�a(ch��n)��(d��ng)�(du��)�S���ص���������ֹ���y(t��ng)�ľS����ȱ���Y��
B.�S��������L(zh��ng)������D(zhu��n)���ʣ�
C.�~���a(b��)�Ӿ����{(di��o)��(ji��)���߹��ܵľS���ء�
Ŀǰ�S���ؠI(y��ng)�B(y��ng)������ҕ��һ��(g��)�P(gu��n)�I���أ������w��(n��i)��(ch��)��ˮƽ
��(y��ng)���܇�(y��n)�p���w��(n��i)֬���Ժ�ˮ���ԾS���صă�(ch��)������?y��n)��~�(d��ng)���@Щ��(ch��)�䅢�c�����ػ��(d��ng)���@����ζ �����ڑ�(y��ng)���l(f��)���ڣ�������̎�������ߵȣ��ι��ˮƽ�ľS���ز���һ����Ч����ˣ��ι�S���ص����c(di��n)��(y��ng) ���ڽ������S��һ�����w��(ch��)���������ڑ�(y��ng)���r(sh��)���o(h��)��(d��ng)�
�F(xi��n)��ˮ�a(ch��n)�B(y��ng)ֳ��ָ�ڸ��ܶȡ����s���l�����_չˮ�a(ch��n)��(d��ng)���B(y��ng)ֳ���ڸ��ܶ�ˮ�a(ch��n)��(d��ng)����B(y��ng)Ҫ���]��
A.�~���r��з��ˮ�a(ch��n)��(d��ng)��ֻ�ܵõ�����������Ȼ�����ϣ���ˣ����Ҫ����Ʒ����еõ������еĠI(y��ng)�B(y��ng)��
B.�~���r��з��ˮ�a(ch��n)��(d��ng)�����ڸ��ܶȼ�Ⱥ����˸���������(y��ng)�������Ү�(d��ng)��Ⱦ���l(f��)���r(sh��)�����������졣�S��ˮ�a(ch��n)��(d��ng)������L(zh��ng)���^(gu��)�ȓ�D�F(xi��n)��׃�ø���ͻ�����Ķ�Ҫ��S�������]��Ҫ����(j��)��ͬ�B(y��ng)ֳģʽ�����L(zh��ng)�l(f��)���A�ζ�������ͬ��ͨ�����ܶ���B(y��ng)Ҫ��߾S����ˮƽ��
3.�S����C�cˮ�a(ch��n)��(d��ng)�����ߵ��P(gu��n)ϵ
�S����C(Vitamin C��VC) ��Ӱ��~���rз����ϵ�y(t��ng)�����I(y��ng)�B(y��ng)��֮һ�������c��(d��ng)���S�������^(gu��)�̣����zԭ�����γɡ��F�x�ӡ�������ͼ��صĴ��x�����ӵ��ܾ����������߷���(y��ng)���~������w��(n��i)�ϳ�VC �����������~����L(zh��ng)�l(f��)���^(gu��)������ه����Դ����(y��ng)��һ�N�������I(y��ng)�B(y��ng)�ء�
��֪VC��һ�N���ЏV���������ܺ��������õ��������ӣ�VC����һ�N�^�õĿ��������{(di��o)��(ji��)��������������ߙC(j��)�w�Ŀ��wˮƽ��Ӱ��خ����wҺ����(y��ng)�⣬߀�ܼ����a(b��)�wϵ�y(t��ng)����(qi��ng)�~�ķ��خ������ߙC(j��)�ܣ�����~�w�����w���������~Ŀ����C(j��)�ư�����棬�б�Ƥ��ճҺ�ӵȱ��o(h��)���ϣ�߀�з��خ��Ժ��خ������߱��o(h��)�C(j��)�ƣ������a(b��)�w��һ�N�dz���Ҫ�ķ��خ����wҺ�������ӡ�
3.1 VC��(du��)�C(j��)�w���ߵ�����ͨ�^(gu��)���Ўחl;���l(f��)�]��
��һ��VCӰ�����(x��)���������ã��Ȯ���(x��)���;��ɼ�(x��)���о����д���VC ��VCȱ�������@Щ��(x��)������������������֮�⣬�������^(gu��)���У���(x��)����(n��i)��VC�M(j��n)һ�����ͣ��a(b��)��VC�ɴ_�����ɼ�(x��)���^�m(x��)�Ƅ�(d��ng)�͵ֿ��������ڑ�(y��ng)����B(t��i)�£��wҺ���ߙC(j��)�����ͣ��a(b��)��VC���@�����خ��ԑ�(y��ng)��Դ��(d��o)�µ����ߑ�(y��ng)�����ã��ڶ���VC����һ�N��������(�����ɻ������)�l(f��)�]�����{(di��o)��(ji��)���ã���?y��n)����ɻ��Ɖļ?x��)��Ĥ��VC���o(h��)�ܰͼ�(x��)��Ĥ������l(f��)��֬�|(zh��)�^(gu��)�������S������ϵ�y(t��ng)�������ԣ�VC�ܺ�������������(����}���ء��S����E)�l(f��)������á�VC��VE���~��еąf(xi��)ͬ�����ѱ��U������ˣ�VCͨ�^(gu��)����VE���m��ˮƽ���^�ߵ�����û��Ժͼӏ�(qi��ng)���������o(h��)���g������(qi��ng)���߷���(y��ng)�������S���،�(du��)ˮ�a(ch��n)��(d��ng)�。��������VC���鿹��������ͨ�^(gu��)�S����Ҫ����(x��)���Y(ji��)��(g��u)���ܵ������Զ�����(qi��ng)���߹���(Chew,1995)��������VC�����I����̴������^(gu��)�����ɶ����M(j��n)���ߣ���?y��n)�̴������������߷��?y��ng)�����ģ�VC����һЩ����ɔ_�غϳɣ���ߙC(j��)�w���߹��ܡ����壬VC��(du��)���߹��ܵ�����(qi��ng)߀������ͨ�^(gu��)���c��(du��)�C(j��)�w�з����C(j��)�ܵĽ���Ԫ���F���~�ȵ��\(y��n)ݔ����(sh��)�F(xi��n)�ġ����⣬VC��(du��)�~���rз���ߵ�Ӱ�߀�Еr(sh��)�g������Ч��(y��ng)����ˣ��M(j��n)һ���о��r(sh��)�g������Ч��(y��ng)�DZ���ġ�
3.2 �S����C�cˮ�a(ch��n)��(d��ng)��ķ��خ�������
3.2.1 �~�����ϵ�y(t��ng)
����ָ�C(j��)�w�R(sh��)�e��������������S���w��(n��i)ƽ��ͷ�(w��n)����һ��(g��)���l(w��i)ϵ�y(t��ng) ,������Ҫ�����Ƿ�����������(w��n)���c���߱O(ji��n)�� 3���档�~�����ϵ�y(t��ng)ָ�~����ߑ�(y��ng)���c�~�w���ټ�
��(x��)��֮�g��һЩ�f(xi��)�{(di��o)ϵ�y(t��ng)�ĽY(ji��)��(g��u)�c�ɷ� ,���~�w��(zh��)�����߹��ܵęC(j��)��(g��u) ,�Ǯa(ch��n)�����ߑ�(y��ng)������|(zh��)���A(ch��) ,���������١�����(x��)�����wҺ�������ӽM�ɡ��~����M(j��n)���������߱��o(h��)�r(sh��) ,��Ҫ��ʹ�خ��Ժͷ��خ��ԃɴ����߷��߹��ܡ��S���،�(du��)ˮ�a(ch��n)��(d��ng)�。���������~���̶��������ڷ��خ��Է���ϵ�y(t��ng)����������ˮ���~���?y��n)��خ������߷��?y��ng)�İl(f��)չ�ܜضȵ�Ӱ푡��خ�������ϵ�y(t��ng)�ͷ��خ�������ϵ�y(t��ng)��ͨ�^(gu��)��(x��)�����ߺ��wҺ���ߙC(j��)�Ɓ�(l��i)�ṩ���o(h��)�Է�ֹ��Ⱦ���������خ��������ɷ�ֹ��ԭ���ֵ��������Ә�(g��u)�ɣ���Ҫ��������������
��(y��ng)�����������ϵK(��Ƥ���[Ƭ��Ӿ��Ƿ�ֹ��ԭ���ֵ��������ϡ����⣬��Ƥ��(x��)�����ڵ�ճҺ��(x��)��Ҳ����Ч�ķ��خ��Է�������)��ø(��Һ�е����^(gu��)������ø��ճҺ��θҺ�c�����M���������е��ܾ�ø)�����ɼ�(x��)��[���ɼ�(x��)��(macrophage)������������(x��)��(Neutrophils)]��ѪҺ����(�ɔ_�ء�C����(y��ng)���ס��a(b��)�w��)��
�������ܲ���ԭ����������\(y��n)��(d��ng)�����Һ������Ʋ�ԭ�������L(zh��ng)�͚��粡ԭ����ijɷ֡������ԭͻ�Ƶ�һ������������Ƥ���~͕�(hu��)���F(xi��n)�ֲ��װY����(y��ng)����Ѫ�����Ѹ��N�c��ԭ����(zh��n)�İ�(x��)��(Leucocytes)�\(y��n)�͵��װY�^(q��)����(x��)���������������ֲ�ԭ�ľ��ɼ�(x��)��(macrophage)���܉��Ɖ��ѱ���ԭ��Ⱦ�ļ�(x��)���ļ�(x��)������(x��)��(cylotoxic cells)����(x��)������(x��)��Ҳ�����ƉĿ�����׃�ɰ���(x��)���Į�����(x��)����
���� (immunity)��һ�N�O���(f��)�s�������F(xi��n)�� ,�ܵ��ܶ����ص�Ӱ� ,�ɸ����鿹ԭ���C(j��)�w�ͭh(hu��n)����������(g��)���� .���~����ߑ�(y��ng)����� ,��ԭ���ذ�����ԭ�����|(zh��)���M(j��n)��C(j��)�w
��;���Լ���ԭ�Ą����� ;�C(j��)�w���ذ����~�����g���w�ء��I(y��ng)�B(y��ng)��r��������B(t��i)�Լ��Ԅe��Ⱥ�wЧ��(y��ng)�� .�@Щ�����L(zh��ng)���ԁ�(l��i)���ܵ��~����ߌW(xu��)�ҵĸ߶���ҕ.Ӱ��~����߰l(f��)��׃���ĭh(hu��n)��������Ҫ�Мضȡ�����(ji��)�����������|(zh��)���ܽ���ˮ�е��ЙC(j��)���o(w��)�C(j��)���ؽ����x�ӵ�
��Ⱦ�� ,�������P(gu��n)�ض��о��Ĉ�(b��o)�����.
�~���c�h(hu��n)���|(zh��)�������������P(gu��n) ,���N�h(hu��n)����Ⱦ�(du��)�~����ߙC(j��)�ܾ��в�ͬ�̶ȵĶ������� ,��(d��o)���~ֿ������������½� .�~��|ˮ�w��Ⱦ��� ,�~�����ϵ�y(t��ng)�ĽY(ji��)��(g��u)�͙C(j��)���܉��^���еذl(f��)��׃�� ,��˿ɑ�(y��ng)���~����߽M�ּ����ܵ�׃������(sh��)��(l��i)�z�y(c��)�h(hu��n)����Ⱦ�ij̶� ,������ɼ�(x��)�����ΑB(t��i)�ͻ���׃������(sh��)�ڭh(hu��n)����Ⱦ�z�y(c��)���ѵõ��V���đ�(y��ng)��.
3.2.2�ך������ϵ�y(t��ng)
�ך�����ߙC(j��)���Է��خ������߷���(y��ng)���������w��(n��i)ȱ������(d��ng)�������е��خ������ߣ����s���R(sh��)�e����������ɷֵķ��خ�������ϵ�y(t��ng)����(du��)�������w��(n��i)�IJ�ԭ������������@�ķ�������(y��ng)���r��з��ؐ�ȟo(w��)����(d��ng)������߷���(y��ng)��Ҫ������(x��)������(cellular immunity)���wҺ����(humoral immunity)��Ѫ��(x��)�����w����(x��)�������w����(x��)��������(x��)�����ڙC(j��)�w��������(y��ng)��������Ҫ�����ã�����Ҫ��ه��Ѫ��(x��)�������ɡ����ҡ��γɼ�(x��)���Y(ji��)�ȼ�(x��)�����߷���(y��ng)����(d��ng)���l����׃������翹ԭ���|(zh��)�̼��r(sh��)���C(j��)�w�w��(n��i)������(x��)��(���ɼ�(x��)��)�l(f��)�]���ã����������ɡ����ҡ��ܽ⡢�ų�����������w��(n��i)���²����������Ȯ���S�������Ľ�����
�wҺ�����������ڟo(w��)����(d��ng)��C(j��)�w�����߷�������(y��ng)�аl(f��)�]ʮ����Ҫ�����ã��@Щ���Ӱ�����Ȼ�γɵĻ��T��(d��o)�a(ch��n)���ĸ��N������Է��ӣ���Ҫ��Ѫ�ܰ��з�����øԭϵ�y(t��ng)��������ø����������ӡ����������ӡ�Ѫ�����ӡ���(x��)���������ӡ��R(sh��)�e���ӡ������ء���Ѫ�ء��ܾ�ø�Լ������ĵȸ��N���������Ե�ø����|(zh��)��
3.3.3 �S����C�����ھS���~�Ƥ�w�ͼך���(d��ng)���⚤�������ԣ�
�~�Ƥ�w�ͼך���(d��ng)���⚤�Ǹ��w������(g��)���w�ⲿ�Ę�(g��u)�죬�c���h(hu��n)���Ľ��|������У��ǵ�����ԭ�w���ֵ���Ҫ�������ϡ�һ��Ƥ�w���⚤�܂������������Ļ��W(xu��)�����S֮��ʧ����(x��)���Ȳ�ԭ�w�͕�(hu��)��̓���������Ⱦ����ˣ���ֹ�~�w���ך���(d��ng)���w�܂������M(j��n)���ڱM�������@�÷dz���Ҫ��
�S����C���M(j��n)�������ϵęC(j��)�ƣ�
�����䅢�c�zԭ���ĺϳɺ���ѿ�M�����γɡ����c�zԭ���ϳ��ǾS����C��Ҫ���������ܣ������zԭ�D(zhu��n)䛷��g������^(gu��)�̣�����������и����ᡢه���Ὓ(j��ng)�u��ø����׃���u��������uه�����^(gu��)���е���Ҫ�o�����ӡ�
ԭ�zԭ�������zԭ����ǰ�w���ǽY(ji��)���M������(chu��ng)�����ϕr(sh��)���۽M���ǻ��|(zh��)�γ��^(gu��)�̱���ġ��S����Cȱ��������r(sh��)��(hu��)��(d��o)�C(j��)�w�zԭ�����u�����㣬�zԭ�������u�����Ậ���@�����ͣ��Ӿ��������ϡ��S����Cȱ����K�M���zԭ������ı��_(d��)��
��ѿ�M�������ǽM���p�����ޏ�(f��)�^(gu��)���г����F(xi��n)�ģ���ë��(x��)Ѫ�܃�(n��i)Ƥ��(x��)���M���w�S��(x��)��������ֳ���γɵģ���������ë��(x��)Ѫ�ܵ����ɽY(ji��)���M�����书���ǵֿ���Ⱦ����������M����Ѫ���K�Ȳ����a(ch��n)������a(b��)�M��ȱ�p���S����C���M(j��n)��ѿ�M�����L(zh��ng)�����䅢�c��ѿ�M����ԭ�zԭ�ĺϳɣ����M(j��n)��ѿ�M����ӵ���������(x��)���;��ɼ�(x��)�����������úͽ���(r��n)(infiltration)���h(hu��n)��(encapsulatory)����(y��ng)���ֿ���Ⱦ�������ԭ�a(ch��n)�����ѿ�M�������L(zh��ng)�߳��ϵK��
3.3.4�S���أÌ�(du��)ˮ�a(ch��n)��(d��ng)���wҺ���ߵ�Ӱ�
�S���أ���һ��������(n��i)��(du��)�wҺ�����д��M(j��n)���� ,���^(gu��)�����㶼�����������á��m���S���أ�Ҳ������(qi��ng)�Ї�(gu��)��(du��)�rѪ�匦(du��)����Ѫ�����Ě������� ,�Ҵ�����(qi��ng)������һ��������(n��i)�ʬF(xi��n)�������P(gu��n)�ԡ���(du��)��ͬˮ�a(ch��n)��(d��ng)���ָ��(bi��o)���� ,�@��������ߵľS���أõ���Ҫ��Ҳ��ͬ���S���о����� ,���C�C(j��)�w������ߠ�B(t��i)�ľS���أ���Ҫ���h(yu��n)���ڙC(j��)�w������L(zh��ng)����Ҫ������(d��ng)�S���أú����� 100mg/kg�D�� (���m���L(zh��ng)������ )�r(sh��) ,��Ӱ����V���w�a(ch��n)�� ;��(d��ng)�S���أü������������m���L(zh��ng)���� 5���� 10���r(sh��) ,�������@���M(j��n)���w�Įa(ch��n)�� ,���� 10���r(sh��)�a(ch��n)����ࡣ
3.3.5 �S���أÌ�(du��)��з���خ������ߵ�Ӱ�
�����c����
��(sh��)�(y��n)��з ���؞�37.52��2.19g�Ľ�����з��ÿһ̎���M10ind.��ÿһ̎����ƽ�У���(sh��)�(y��n)����60d��
��(sh��)�(y��n)�O(sh��)Ӌ(j��)��
����(sh��)�(y��n)̎���M�� ���1�H����A(ch��)��ϣ����2����A(ch��)��ϣ�250mgVC�����3����A(ch��)��ϣ�500mgVC�����4����A(ch��)��ϣ�1000mgVC�����5����A(ch��)��ϣ�1500mgVC��
�� ��(j��)* ������ø(PO) ��������(Ua)���ܾ�ø (UL)�����������绯ø(SOD)���A������ø(ALP)����������ø(ACP)
3.3.5 �S���أÌ�(du��)��з���خ������ߵ�Ӱ�
�Y(ji��)��1�� �S����C��(du��)��зѪ����PO���Ե�Ӱ�
�Y(ji��)��2���S����C��(du��)��зѪ�塢����������п�������Ӱ�
�Y(ji��) ��3�� �S����C��(du��)��з���M�����ܾ�ø������Ӱ�
�Y(ji��) �� 4���S����C��(du��)��з���M����SOD������Ӱ�
�Y(ji��) �� 5�� �S����C��(du��)��з���M����ALP���Ե�Ӱ�
�Y(ji��) �� 6�� �S����C��(du��)��з���M����ACP���Ե�Ӱ�
С �Y(ji��)
3.3.5.1 �S����C��һ����õ����ߴ̼������m����������Ч��ߺ�з���M����PO��Ua��UL��ALP��ACP�ȵĻ��ԣ��Ķ�����(qi��ng)��з�ķ��خ�����������
3.3.5.2 ����Ч����(qi��ng)��з���خ�����������VC���m���������քe��500mg-1000mg/100g��ϡ�
�S����C����a(b��)�w(complement body)�����a(b��)�wϵ�y(t��ng)���~ֿ������Ⱦ����Ҫ�ɷ֣��ɴ������wҺ�еĔ�(sh��)ʮ�N����ø���Ե��M�ɡ��~��a(b��)�w���c�خ��Ժͷ��خ��Է����������ɽ�(j��ng)��;��(classical pathway)����·;��(alternative pathway)����ļ�(x��)���ܽ����ú��ɱ������a(b��)�w�M��ጷŵ�Ƭ������ʹ���{(di��o)������(oponization)
VC��(du��)�a(b��)�w�Ŀ������ÙC(j��)���ǣ����a(b��)�w�Ľ�(j��ng)����(d��ng);���У�C1��(f��)���w(C1q��r��s)���ȱ����w�����VC���܌�(du��)C1��(f��)���w��Ӱ푡�C1q�и����u��������zԭ���טӅ^(q��)��VC�t���c�u��������u��������.
�S����Cȱ�������~��a(b��)�w���ԣ��ܿ�����������K�˸������ه�����u������(y��ng)��ɵġ��߄����ľS����C����a(b��)�wC3��ˮƽ���Ķ�����a(b��)�w�Ļ��ԡ�
3.3.5 �S���أÌ�(du��)ˮ�a(ch��n)��(d��ng)�:(x��)�����ߵ�Ӱ�
3.3.5.1�S����C������ɼ�(x��)�������������͚��������ɼ�(x��)�����~��خ��Լ�(x��)����������Ҫ��(x��)�����~�����(x��)�����κ˼�(x��)���;��ɼ�(x��)�������������á������ɚ����^(gu��)�̰�����څ���c�R(sh��)�e���������ɰ�����������Ͱ����ų��Ă�(g��)���E������(x��)�����κ˼�(x��)�����~�����Ѫ�д������ڣ����ɼ�(x��)�����~��I��Ƣ���ܰͼ�(x��)���к����S�����߄����ľS����C����ߺ��V�������Һ������[�������ɼ�(x��)�������ɻ��ԣ�������ߺ��V���ɼ�(x��)���İ���ԡ�
3.3.5.2�S����C������ɼ�(x��)�������ɻ��ԵĿ��ܙC(j��)�ƣ�
A.������ɼ�(x��)�����\(y��n)��(d��ng)څ���ԣ��߄����ľS����C�����������������(x��)�������ߺ͚������ܣ�߀������߭h(hu��n)�B�����ǰ������EI�ĝ�ȣ���ֹ���ɼ�(x��)���ļ�(x��)��Ĥ���������Ķ�ʹ��(x��)����څ���Ե��ԾS�֡�
B.Ӱ����ɼ�(x��)��څ�����ӵĺϳɣ��a(b��)�w����C5a��Ó������C5a�����ɼ�(x��)��څ�����Ӽ�(x��)������څ�����á��S����C���w��(n��i)��߀ԭ�����|(zh��)���f���w�����c�w��(n��i)���N����߀ԭ����(y��ng)������Ӱ��ض���ø����(y��ng)�����ɼ�(x��)��څ�����ӵĺϳɣ��Ķ�Ӱ����ɼ�(x��)�������ɻ��ԡ�
3.3.5.3 �S����C��(du��)ˮ�a(ch��n)��(d��ng)�:(x��)�����ߵ��о��Y(ji��)��
�D���м���S���أõ����� 1000�� 4000mg/kg�r(sh��) ,���@����ߺ��V���ɼ�(x��)���Ļ���������ʯ���~�D���оS���أ��������� 2000mg/kg�r(sh��) ,��ѪҺ��(x��)������(sh��)����ͬ�N�(x��)�� (�����ܰͼ�(x��)������������(x��)���͆κ˼�(x��)�� )�M�ɱ����s���ܾS���أõ�Ӱ푡��@�f(shu��)���S���أÌ�(du��)���خ��Լ�(x��)�����ߵ�Ӱ� ,���H�c�S���أõ������P(gu��n) ,����߀�܂�(g��)�w���Ӱ푡����ҾS���أÌ�(du��)���خ��Լ�(x��)�����ߵĴ��M(j��n)Ӱ푿��ܴ���һ��(g��)�ֵ ,�S���أ��ڴ��ֵ������(n��i)��������˺̈́�(d��ng)��ķ��خ��Լ�(x��)�����߹��� ,���ڻ���ڴ��ֵ�������������� ,����ͬ�Ă�(g��)�w��ͬһ��(g��)�w�ڲ�ͬ�ėl�������ֵ��ͬ��
�P(gu��n)�ھS���أÌ�(du��)���ɼ�(x��)���������õ�Ӱ� ,���о��Y(ji��)����������֮�g�o(w��)���@���P(gu��n)ϵ�������T��(du��)�۵��A�������ɺ͚������ò��ܾS���أõ�Ӱ� (Li�� ,1985)���D�ϾS���أÌ�(du��)�������q�^�I���ɼ�(x��)������SRBC��a(ch��n)������������x�� (��2 -)�������o(w��)���@���� (Hardie�� ,1991 )���D���оS���أõ������� 30�� 3000mg/kg�r(sh��) ,��(du��)���c(di��n)��β�~�صİ�(x��)���������Û]��Ӱ푡�
��ʯ���~�ھS���أ��������� 2000mg/kg�r(sh��) ,���خ��Լ�(x��)�������(d��ng) (����ѪҺ��(x��)�����^�I��(x��)�������ɻ��(d��ng) )Ҳ�]���ܵ�Ӱ푡���Ҳ�ЌW(xu��)�߈�(b��o)�� ,�߾S���أ�(3000mg/kg )�������D�������@����(qi��ng)���V���ɼ�(x��)�������ɻ���;�ø߾S���أú����D���ι�Ľ��^�� ,�����ɼ�(x��)�����ɻ����Ȍ�(du��)�սM���@�����;�m���S���أ���һ��������(n��i)�ʬF(xi��n)�������P(gu��n)�Ե�����(qi��ng)��(du��)�rѪ��(x��)����(du��)���Sɫ����������������ü������������ ,��(du��)�ھS���أÌ�(du��)�������õ�Ӱ� ,߀��Ҫ�M(j��n)һ���о��C��(sh��)��
3.3.6 �S����C��(du��)�خ������ߵ�Ӱ�
3.3.6.1�~��خ�������
�خ���������ָ?j��ng)�(g��)�w�ڳ�����������^(gu��)���У����|��ԭ�������γɵČ�(du��)��ԭ������ᘌ�(du��)�Ե������������c����~��خ������ߑ�(y��ng)��(y��ng)����Ҫ�M�����������I�K�����١�Ƣ�K�������� �ܰͽM����ѪҺ�ܰͽM������(x��)����Ҫ�����ɼ�(x��)�����ܰͼ�(x��)���������ף���Ҫ��IgM���Լ�һЩ�wҺ�ܰ����ӵȡ�
�~Ŀ��w�a(ch��n)�����������c(di��n)��
�ٿ��w�γ����^�L(zh��ng)�����w�ζ������^������ˮ���~t���L(zh��ng)��
���ڳ��Α�(y��ng)���У��~��w���m(x��)���^�L(zh��ng)��
������ӛ��������ˮ�ص�Ӱ푣����Α�(y��ng)���c���Α�(y��ng)���w�r(ji��)�ı���һ���^�ͣ��е��~��o(w��)�^(q��)�e���״����ߺ��ٴ����߮a(ch��n)���Ŀ��w����IgM��
3.3.6.2 �S����C��(du��)�~��خ������ߵ��о���
�S����C�����ߑ�(y��ng)����������Ҫ�����á��߄����ľS����C�����Դ��M(j��n)���V�ʹ������qT�ܰͼ�(x��)������ֳ�����������B(y��ng)��(sh��)�(y��n)��(j��ng)�c���o�S����C�����VT�ܰͼ�(x��)������ֳ��ߡ��о�߀�l(f��)�F(xi��n)���߄����ľS����C��߀����ߺ��VB-�ܰͼ�(x��)������ֳ�����⣬�߄����ľS����C�ܴ��M(j��n)�~�(du��)ijЩ��(x��)���ԺͲ����Բ�ԭ�w���خ��Կ��w�a(ch��n)������߄����ľS����C������ߺ��V�����c(di��n)��β�t�ʹ������q�خ��Կ��w�Įa(ch��n)����
3.3.6.3�S����C���M(j��n)�ܰͼ�(x��)����ֳ���خ��Կ��w
�a(ch��n)���Ŀ��ܙC(j��)����
�پS����Cʹ�ܰͼ�(x��)�����������p��������ܰͼ�(x��)������ֳ���خ��Կ��w�Įa(ch��n)�����ܰͼ�(x��)�������߷���(y��ng)�����ģ���DNA���ܵ����N���|(zh��)������������ɼ�(x��)������������(x��)�����װY����(y��ng)����T-�ܰͼ�(x��)���ēp����
�� �S����C���c���װ���������ζ����ķ���(y��ng)���ڿ��w�ĺϳɡ��خ��Կ��w�����S������I�������Ƀɂ�(g��)���װ����B�Ӷ��ɵġ������װ��������װ������ɵģ��η���(y��ng)��Ҫ�S����C �ą��c��
�� �����ǽ�����һЩ�ؽ��ٌ�(du��)�~�����ϵ�y(t��ng)�a(ch��n)���Ķ���Ӱ푣����ж������x���D(zhu��n)����߀ԭ�B(t��i)��
3.3.7 �S����C�����ˮ�a(ch��n)��(d��ng)���(y��ng)��������
�S���أ�������(qi��ng)�~���r��з��ˮ�a(ch��n)��(d��ng)�(du��)���N��(y��ng)�����m��(y��ng)������Ishibashi(1992 )��(b��o)��,�m���ľS���أ�������(qi��ng)��������(y��ng)�������������c(1996)��(b��o)��,�m���ľS���أ�����Ч����Ї�(gu��)��(du��)�r��ȱ����������Merchie(1995)�l(f��)�F(xi��n),�ߝ�ȾS���أ����B(y��ng)��Artemia nauplii�ι���c(di��n)��β�t���� ,������(qi��ng)��(du��)�}�ȵĵֿ�����������ȣ�2001���о��l(f��)�F(xi��n)����������Ӹߺ����ľS����C��������A�q��з�������w����з�Ŀ���(y��ng)��������
3.3.7 Ӱ푾S����C����~� ���ߙC(j��)�ܵ����أ�
�� �S����C����ʽ��
�ھS����C�Ą�������(sh��)�(y��n)�����܌����ӄ����c���a(ch��n)���ܡ�Ѫ�{�������M�������оS����C�ĝ��׃���Y(ji��)������(l��i)�����S����C�Č�(sh��)�H��Ч�������ܕ�(hu��)����(zh��n)�_�����⣬��ͬ�����߅���(sh��)����(du��)�S����C��������Ҳ��ͬ��
�۾S����C�����Õr(sh��)�g�����r(sh��)Ч�P(gu��n)ϵ��
�� �h(hu��n)���l������ض�
3.8 �S����E��(du��)ˮ�a(ch��n)��(d��ng)�����ߵ�Ӱ�
�S����E(Vitamin E��VE)��(du��)ˮ����(d��ng)�����߹��ܵ�Ӱ��ǬF(xi��n)���о��ğ��c(di��n)���@������о��ɞ�ˮ����(d��ng)�R���ķ����ṩ�I(y��ng)�B(y��ng)�W(xu��)�����֧�֡���(d��ng)�����߷���(y��ng)����(d��ng)��Ҫ�ڼ�(x��)��Ĥˮƽ�l(f��)���������w֬��Ҫ����������VE�Ǽ�(x��)��Ĥ��һ��(g��)��Ҫ�ɷ֣�����ϵ�y(t��ng)��f(xi��)������²��ЙC(j��)����a(ch��n)���IJ���(w��n)�����ɻ���(hu��)�Ɖ����ɼ�(x��)������VE�ɱ��o(h��)�w��(x��)���������ɻ��ƉIJ�ʹ��(x��)��Ĥ�S����������(Tengerdy et al.��1981)�����߱��o(h��)�����cǰ������ˮƽ�������P(gu��n)��E��ǰ
�����ظɔ_����ϵ�y(t��ng)�Ĺ��ܣ��T���ܰͼ�(x��)���Ļ������ܰͼ�(x��)������ֳ�Լ����ɼ�(x��)����һϵ�й��ܡ�
VEͨ�^(gu��)����ǰ�����غ�Ƥ�|(zh��)ͪ������ϳɣ����M(j��n)�wҺ�� ��(x��)�����ߺͼ�(x��)�����������Լ����IL��I������(l��i)����(qi��ng)�C(j��)�w���߹��ܡ���?y��n)�ǰ�����������߷��?y��ng)�͙C(j��)�w���������wϵ��Ҫ�{(di��o)��(ji��)����ɔ_T�ܰͼ�(x��)�����Ժ��нz����ԭ�錧(d��o)�ܰͼ�(x��)����ֳ����������߿ɮa(ch��n)���������ƣ�Ƥ�|(zh��)ͪ�������ߡ� VE�ɷ�ֹ���Ⱦ���T�l(f��)��ǰ�����غ���������
(Shetty et al.��1979��Likoff et al.��1981)��
���B(y��ng)ֳ�~���r��з�ˮ�a(ch��n)��(d��ng)��������a(b��)��VE�ܾS�ֲ�����(qi��ng)�~���r��з�������(x��)�����wҺ���߹��ܣ�����~���r��зĿ�����(Tengerdy��1990)��������a(b��)��VE�ܴ��M(j��n)���ɼ�(x��)����������������߰��c(di��n)��β�t�ʹ������~���ɼ�(x��)�������ɻ�����Wise et al.��1993��Pulsford et al., 1995 ��������(qi��ng)���V��(du��)��(x��)����ԭ�w����������Wise et al.��1993��,���M(j��n)���V��Y. ruckeri�Ŀ��w�a(ch��n)��(Ndoye et al., 1990)�����ܶȗl���£�����VE�܉�����a(b��)�w���ԡ�Ȼ�� ����и�ˮƽ��VE����������(qi��ng)�������qѪ�����a(b��)�w������Ҳ��Ӱ��wҺ���w����(y��ng)(Lall et al., 1988; Hardie et al.,1990)��
������a(b��)��VE���@������(qi��ng)�Ї�(gu��)��(du��)�rѪ���з�����ø��PO�������������S��VE��ȵ�����PO��������(qi��ng)���m����VE�ܴ��M(j��n)������øԭ(proPO)�ĺϳɣ�����(qi��ng)�Ї�(gu��)��(du��)�rѪ�匦(du��)����Ѫ���������廡�������ɻ����������c��1996�����S�����ɼ�(x��)�����ɻ��������VE��ˮƽ��ه�ڌ�(du��)��ԭ�w���R(sh��)�e�̓�(n��i)�ݠ�r�����������gVE��Ҫ�ǾS�ּ�(x��)��Ĥ�Ĺ��ܣ�Tengerdy et al.��1984����VE���M(j��n)�z��ԭ�T��(d��o)ƢT��(x��)������ֳ������(qi��ng)������A(ConA)�Ĵ��ܰͼ�(x��)����ֳ���ã������ܷ�ֹ֬���ጦ(du��)�ܰͼ�(x��)����ֳ���������á�
VE�������ܰͼ�(x��)�����e���o����T�ܰͼ�(x��)����ֳ��T��B�ܰͼ�(x��)���f(xi��)ͬ���ü����ɼ�(x��)����ֳ��VE���H���鼚(x��)����(n��i)����������(w��n)����֬���ἰ֬���ϳ��c�ֽ�����g�a(ch��n)�ﲻ�������Ɖģ�����Ӱ푻�����ϩ��Ĵ��x��ǰ�����صĹ��ܡ�VE��(du��)���ߵĴ��M(j��n)���þ������@�Ą���Ч��(y��ng)�P(gu��n)ϵ������������ߑ�(y��ng)������VE�h(yu��n)�h(yu��n)���^(gu��)������L(zh��ng)��(du��)VE����Ҫ���ڰl(f��)����Ⱦ���r(sh��)������VE�����ø���
3.9 �S����E��(du��)��з���خ������ߵ�Ӱ�
�����c����
��(sh��)�(y��n)��з ���؞�37.52��2.19g�Ľ�����з��ÿһ̎���M10ind.��ÿһ̎����ƽ�У���(sh��)�(y��n)����60d��
��(sh��)�(y��n)�O(sh��)Ӌ(j��)��
����(sh��)�(y��n)̎���M�� ���1�H����A(ch��)��ϣ����2����A(ch��)��ϣ�10mgVE�����3����A(ch��)��ϣ�20mgVE�����4����A(ch��)��ϣ�40mgVE�����5����A(ch��)��ϣ�60mgVE��
�� ��(j��)* ������ø(PO) ��������(Ua)���ܾ�ø (UL)�����������绯ø(SOD)���A������ø(ALP)����������ø(ACP)
3.9 �S����E��(du��)��з���خ������ߵ�Ӱ�
�Y(ji��)��1�� �S����E��(du��)��зѪ����PO���Ե�Ӱ�
�Y(ji��) �� 2�� �S����E��(du��)��з���M���п�������Ӱ�
�Y(ji��) �� 3�� �S����E��(du��)��з���M�����ܾ�ø������Ӱ�
�Y(ji��) �� 4�� �S����E��(du��)��з���M���г��������绯ø���Ե�Ӱ�
�Y(ji��) �� 5�� �S����E��(du��)��з���M���ЉA������ø���Ե�Ӱ�
�Y(ji��) �� 6�� �S����E��(du��)��з���M����ACP���Ե�Ӱ�
ӑՓ
A. PO�Է�����øԭ(proPO)����ʽ�������w����(x��)���У�������øԭϵ�y(t��ng)
(proPO System)�ڼך���(d��ng)�������R(sh��)�e�ͷ������ã�proPO����ڼ���(d��ng)����a(b��)�wϵ�y(t��ng)���Ǽך���(d��ng)���wҺ���ߵ���Ҫ�M��, �����������D(zhu��n)������л��Ե�
PO (Soderhall, 1982; Ashida et al, 1982; Johansson et al., 1989)��POһ���慢�c�ڻ���������(y��ng)�к�ɫ�صĺϳ�(Aspan et al,1991)����ɫ�ؿ������Ʋ�ԭ�w���⵰��ø�͎��|(zh��)ø�Ļ��ԣ��Ķ��ڂ������ϡ���������������ԭ�w����l(f��)�]����Ҫ�����ã���һ��������һ�N�R(sh��)�eϵ�y(t��ng)�T��(d��o)��������ϵ�y(t��ng)�������ɷֵĮa(ch��n)�����Ɯy(c��)VE����ߺ�зѪ����PO���ԵęC(j��)���ǣ���������Ȼ������������ܾS�֙C(j��)�w�����������ܣ��ӿ��з��ꐴ��x���m����VE�ܴ��M(j��n)������øԭ(proPO)������ϳɣ����������øԭ(proPO)����ʹproPO�D(zhu��n)����PO���Ķ����Ѫ����PO���ԡ��M�����������VE����ߺ�зѪ����PO���ԣ������������зѪ��(x��)����proPO������(qi��ng)PO���Ե����ÙC(j��)�ơ��Լ���ͬ�l����VE����Ч�������в�������д�������ϵ�y(t��ng)���о���
ӑՓ
��(sh��)�(y��n)�Y(ji��)��������VE��Ӱ푺�з���M���п��������ܾ�ø��������һ��������(n��i)���S��VE���������ߣ���з����(y��ng)�M���п��������ܾ�ø����Ҳ�S֮����(qi��ng)��VE�����^(gu��)60mg/100g��ϕr(sh��)�����M���п��������ܾ�ø�����������͡��ɴ˿�Ҋ���m������VE����ߺ�з���M���еĿ��������ܾ�ø����������(qi��ng)��з�ķ��خ������������Ķ���ߺ�з�Ŀ�����������VE����ߺ�з���M���еĿ��������ܾ�ø�����ęC(j��)���ǣ����������з���L(zh��ng)�l(f��)���^(gu��)���еı���I(y��ng)�B(y��ng)�أ���ʹ�C(j��)�w̎��һ�N���õ�������B(t��i)������Ч�ش��M(j��n)�C(j��)�w����ꐴ��x���{(di��o)��(ji��)�C(j��)�w��������������(y��ng)�����M(j��n)�������ܾ���������ϳɣ��Ķ���ߙC(j��)�w�Ŀ��������ܾ�ø����������(qi��ng)�C(j��)�w�Ŀ�������
��(d��ng)��C(j��)�w��(n��i)�ɿ�����øϵ�y(t��ng)�Ϳ��������|(zh��)�M�ɵĿ������wϵ����������w��(n��i)��������ɻ���ʹ���ɻ��a(ch��n)���c����̎��һ�N��(d��ng)�B(t��i)ƽ���ЏĶ�������ɻ���(du��)������ӵēp���ȡ��о��l(f��)�F(xi��n)��SOD�����c���������ˮƽ�������P(gu��n)����ˣ�SOD������C(j��)�w���خ�������ָ��(bi��o)�����Á�(l��i)�u(p��ng)�����ߴ̼�����(du��)�C(j��)�w���خ�����������Ӱ푡�
��з���������VE��(hu��)�@������������(y��ng)�M����SOD���ԣ��@��Ҫ��VE�����l(f��)�]�˿��������õĽY(ji��)�������������20mg-40mg/100g���VE�r(sh��)����з���M����SOD���Ծ����ͣ��ҽ������Ğ�����٣�������ѳ����@����������VE���ڸ����ٺ��ѳ��зe�ۣ����l(f��)�]�����������������õĽY(ji��)�������̈́���̎���M����Ͷι���������10mg/100g���VE�ĺ�з���H������SOD�����p��׃�����@��������������е�VE�����^�ͣ���Ҫ���������L(zh��ng)�l(f��)�����^�ٷe�ۡ����⣬�IJ�ͬ�M����SOD����׃������Ҳ���Եó���VE�ں�з�C(j��)�w��(n��i)�ķֲ��Dz�����ģ���Ҫ�ֲ���֬�������^��ĽM�����٣�������ٺͿ��ٰl(f��)�����ѳ�.
��(sh��)�(y��n)�Y(ji��)���������m������VE������(qi��ng)��з����(y��ng)�M����ALP���ԣ������m�����ӷ�����(n��i)���SVE�����������ӣ�ALP��������(qi��ng)��ALP��ֱ�Ӆ��c�Ĵ��x�����cDNA��RNA�������|(zh��)��֬�|(zh��)�ȴ��x���P(gu��n)������(du��)�}�|(zh��)����ȡ�������}���e�������γɡ��ך��ط��ڼ��γɶ�����Ҫ
�����ã��rзȼך���(d��ng)�������L(zh��ng)�^(gu��)���ж�Ҫ��(j��ng)�v͑���^(gu��)�̣�ԓø��(du��)�rз����桢���L(zh��ng)���e��Ҫ�����x��ALP���Ե����ߣ��ɼ��ٺ�з��(x��)���е����|(zh��)���x����ADP���ữ�γ�ATP�ṩ��������Ҫ�ğo(w��)�C(j��)�ף��Ķ����M(j��n)��з���L(zh��ng)��ͬ�r(sh��)Ҳ�g�ӵ�����(qi��ng)������خ�����������������(d��ng)VE���������60mg/100g��ϕr(sh��)����з����(y��ng)�M����ALP���Է����½��ˣ��@�Ƿ������������VE�����^(gu��)�࣬�����ɔ_�˙C(j��)�w��������ꐴ��x���Ķ�����ALP���ԣ��д��������о��� �S���،�(du��)ˮ�a(ch��n)��(d��ng)�。��������
ACP���w��(n��i)Ҳֱ�Ӆ��c������F(tu��n)���D(zhu��n)�ƺʹ��x�����������VEҲ������(qi��ng)��з���M����ACP���ԣ����ʬF(xi��n)���S��VE�������ӣ�ACP����������څ��(sh��)��ACP������ߣ��C(j��)�w�Ĵ��x������ʢ��������(qi��ng)�C(j��)�w�w�|(zh��)���Ķ��g������(qi��ng)��з���خ�������������
3.9 �S����E��(du��)��з���خ������ߵ�Ӱ�(�Y(ji��)Փ)
������ø(PO)��������(Antibacterial activity)���ܾ�ø(Lysozyme)�����������绯ø(SOD)���A������ø(ALP)����������ø(ACP)��ø�ڼך��ѪҺ�ͼ�(x��)���Г�(d��n)ؓ(f��)���C(j��)�w��������Ҫ���ܣ�����VE����з���M����PO��UL��Ua��ALP��ACP�����@����ߣ���SOD�������@���ͣ��f(shu��)���@�ɷN�S�����ں�з�w��(n��i)���F(xi��n)�����^��(qi��ng)�Ŀ������ԣ��ڴ��M(j��n)��з�������L(zh��ng)��ͬ�r(sh��)��Ҳ��Ч������(qi��ng)�˺�з���خ������������������P(gu��n)PO��UL��Ua��SOD��ALP��ACP���ں�з���߷�������(y��ng)�еĵ�λ�����w���ü��S�������T��(d��o)�a(ch��n)���ęC(j��)����������ϵ�y(t��ng)���о�������(sh��)�(y��n)�Y(ji��)����ʾ�� VE���������з����Ч�����ߴ̼��������a(ch��n)��(sh��)�`�м��ԑ�(y��ng)�á��C�Ϸ����ó�����ÿ100g���������VE20��40mg�r(sh��)����Ч������(qi��ng)��з�ķ��خ��������������Ķ���ߺ�з�Ŀ�������
4 .0 �S����B6��(du��)ˮ�a(ch��n)��(d��ng)�����ߵ�Ӱ�
4.0.1 �S����B6
�S����B6�����ߴ�������ȩ�����߰����N���������Ľy(t��ng)�Q�����������ߴ�������ࡣ�S����B6�ǟo(w��)ɫ������ˮ�ʹ��ľ��w����(du��)�ᡢ��͉A��(w��n)������(du��)�����ж����Ɖġ�����Ҫ����������ȩ����ʽ�����oø���c��(d��ng)���w��(n��i)���S����x����(y��ng)��������������x��̼ˮ��������x��֬�����x����Ҫͨ�^(gu��)�D(zhu��n)�������á��o(w��)����Ó�������á�Ó�Ȼ����úͰ������Ó�����ö��l(f��)�]���á�
4.0.2�S����B6�c��(d��ng)�R�����P(gu��n)ϵ
��(d��ng)�����ܵ���Ⱦ���w��(n��i)������ϵ�y(t��ng)�l(f��)������(y��ng)���ϳ��µĵ����|(zh��)������ɼ�(x��)�����ɔ_�غͿ��w�ȡ��@����ζ��Ӱ푵����|(zh��)���x���κΠI(y��ng)�B(y��ng)ȱ�ݻ�ƽ�⣬����(hu��)��(du��)�C(j��)�w�����߷���(y��ng)��ɇ�(y��n)�صēp�����S����B6�ڵ����|(zh��)���x�������P(gu��n)�I�����á�ˮ�a(ch��n)��(d��ng)�(du��)�ռZ�еĵ����|(zh��)�����^�ߣ���������ʳ��ˮ�a(ch��n)��(d��ng)�������(y��ng)�،�(du��)�S����B6�Ĵ��x������Ҳ�^�ߣ��S����B6��ȱ�����C��(sh��)��(du��)���߹����Гp�����á�
4.0.3�S����B6�c��(d��ng)�R�����P(gu��n)ϵ
���~��F(xi��n)��ؚѪ����ʳ���wɫ׃�ڡ�ʧȥƽ�⡢���L(zh��ng)������������;�������q�ľS����B6 ȱ���Y�����������ߡ��О鮐�����I���ѳ��ε���׃���נ��ٜp�١��I����Ѫ�M������������ ,���c(di��n)��β�t����(j��ng)ʧ�{(di��o)���鴤�������ʸߡ��w���{(l��n)ɫ�İY�� ;�ձ����~���F(xi��n)�����L(zh��ng)��������ʳ���d�B���@�� ;�q�V���F(xi��n)��d�B���@�ʡ��̼��^(gu��)�����������(d��ng)���������١��w�w�����ȡ��r�S����B6 ȱ���Y�����ؾ���������ʽ��͡��zʳ�p�١�
1978��Harver��Barnon�ڴ��״�����~��ͬ�����|(zh��)ˮƽ���ռZ�У���׃�S����B6�ĺ�������ʹ�䱩¶��Vibrio angularium�ĭh(hu��n)���У���(sh��)�(y��n)�l(f��)�F(xi��n)�ι��ˮƽ�����|(zh��)�ռZ���~Ⱥ�����ʸߣ����ҾS����B6��(du��)���״�����~����������Ч��(y��ng)��Ҋ�D�������@����Կ������ռZ���m���ľS����ˮƽ�dz���Ҫ����?y��n)��m���ľS����ˮƽ�����H���ԝM��C(j��)�w�Ĵ��x��Ҫ�����ҿ��Ա��C�C(j��)�w�l(f��)�]���������߹��ܡ�
4 .1�S���أ�
֬���Ե�VA��(du��)��Ƥ��ճҺ�ӵ��������Լ�ճҺ�ķ��ں���Ҫ,�ɽ����~�w��(du��)һЩ��Ⱦ�Լ���������,��(qi��ng)�����ɼ�(x��)���Ĺ��ܣ�Blazer,1992�����ԣ��������L(zh��ng)��������VA�ι�q�~�Ͱ��c(di��n)��β�t,���@���p������c(di��n)�x���������������Thompson�ȣ�1994������������Ӹ߄�����VA��ʹ
�������q�������������,��(du��)A. almonicid�ĵֿ������ӡ� �S���،�(du��)ˮ�a(ch��n)��(d��ng)�。��������
4 .2 �r���،�(du��)ˮ�a(ch��n)��(d��ng)�����ߵ�Ӱ�
4.2.1�r����
�r������һ�N�tɫ����}���أ��V���Ĵ�������Ȼ�磬�����Ǻ���h(hu��n)���С��r�����ѱ��C�����ж�N������������ܣ������@Щ�������ܾ��c�俹�������������P(gu��n)��
�о��������r������һ�N�ǾS���أ�Դ����}���� ,�ڄ�(d��ng)���w��(n��i)�����D(zhu��n)׃?y��u)�S���أ�,��(d��ng)�(sh��)�(y��n)���� ,�r���ؿ������NO2 ����������� ,Ҳ�ɽ���֬�|(zh��)�^(gu��)�������� ,��Ч���������ɻ����l(f��)��֬�|(zh��)�^(gu��)�������r���ؾ��Џ�(qi��ng)�ҵĿ��������ԣ�����������}���ظߣ�0�����ȾS���أŸ�550�������Q�顰����(j��)�S���أš��������п����Y����˥�ϡ�����(qi��ng)����������Ҫ���������ܣ����Ҍ�(du��)���w�^��(du��)��ȫ��
4.2.2 �r���ص���ȡ
�r���ؿ��˹��ϳɺ��������w��(n��i)�ϳɡ��˹��ϳɵ��r���ش�����ʽ�Y(ji��)��(g��u)�������w�ϳɵ��r���ش���鷴ʽ�Y(ji��)��(g��u)����(d��ng)���w��(du��)�˹��ϳɵ��r�������������^���������˹��ϳɵ��r���ص���ɫ����������Ч�r(ji��)�h(yu��n)����Ȼ�r���صͣ����Ŀǰ���r�������a(ch��n)��Ҫ�Ǐ�ˮ�a(ch��n)Ʒ�U����ʹ�Ҏ(gu��)ģ���a(ch��n)�ΰ��塢��ĸ����ȡ��Ȼ�r���ء�
(1). ��ˮ�a(ch��n)Ʒ�U��������ȡ�r����
���� ,�яU���������p����ϩ���� ,����-70�汣��,�Õr(sh��)����ɸ���� ,�������r�ַۡ��������� 1�� 1�Ӵ����ڷ�������r�� ,�������,�����U�K������������(l��i)�ܹ⡣40�� 50��ӟ�,�����?c��i)��?���ӟᵽ 90��ֹͣ ,���õ͜��x�� (0��, 11000 r/min,10),
�ռ��r���ص�����Һ�ֳ��p����ό���Һ,�÷�Һ©�����x,ӛ�ɫ����Һ���w�e ,�÷ֹ���Ӌ(j��)��(l��i)����ɫ������ȡ��Ĺ��V����,�� 485nm̎�y(c��)���������ա��r��������Һ������ϵ��(sh��) E�� 1.55,��
����Ĺ�ʽӋ(j��)���r���ص���:
�r���� (��g/ g�U���� )=AD/ 100��GdE
ʽ�� :A:����L(zh��ng)̎������ֵ;D:���յ����w�e(ml)��ϡጱ���(sh��);G:���r�U���� (mg);d:��(x��)������ (1cm);E:����ϵ��(sh��)��
(2) ����������a(ch��n)�r����
Ŀǰ��(b��o)�������a(ch��n)�r���ص����������t���塢�����t���塣�����t�������r���غ������_(d��)10 - 40mg.g-1; ����Ȼ������Ȼ�r���غ�����ߵ�������ô�Ҏ(gu��)ģ�B(y��ng)ֳ�t�������a(ch��n)�r������Ŀǰ��Ȼ�r���ع��I(y��)�����a(ch��n)��һ�l��Ҫ;�������üt�������a(ch��n)�r����һ��փɂ�(g��)�A�Σ����������m���L(zh��ng)�l�������B(y��ng)�t����Ą�(d��ng)��(x��)���@ȡ����ȵ���������Ȼ��ͨ�^(gu��)��׃���B(y��ng)�l���T��(d��o)�r���صĴ����e�ۡ�
(3)���ý�ĸ���a(ch��n)�r����
��(gu��)����Ҫ���üt�l(f��)���ĸ���a(ch��n)�r���ء������Ǻ��w�S���Ǟ��ĸ���e���r���ص��^��̼Դ����ĸ��͵����˞��^�õ�Դ���m���������t��֭�����ם{����ϩ����ȶ�����߽�ĸ���r���صĺ������ߵij�ʼ̼����Ҳ�������r���خa(ch��n)������ߡ�
4.2.3 �r������ˮ�a(ch��n)�ϵđ�(y��ng)��
�r��������һ�N�I(y��ng)�B(y��ng)�ɷ֣���һ���̶��Ͽɱ����֞�S�������ˮ�a(ch��n)�B(y��ng)ֳ�I(y��)�У��r���ؾ��������B(y��ng)ֳ��(du��)����ɫ(�粻ͬƷ�N���r�w��(n��i)�r�����c�����|(zh��)�Y(ji��)�Ϻ���γɸ��N�خ����ɫ�����{(l��n)ɫ���Gɫ����ɫ��)����ߴ����(�r���ؿ��ԅ��c�~���r�w��(n��i)�����wϵ���{(di��o)��(ji��)�������ڂ�(g��)�w�Ĵ��ͽ�����r�ĸ���)�����M(j��n)���L(zh��ng)����ֳ�Ͱl(f��)������Ҫ���á��S��ȫ��ˮ�a(ch��n)Ʒ���������������ӣ��r������ˮ�a(ch��n)�B(y��ng)ֳ�I(y��)�е��Ј�(ch��ng)Ҳ�������U(ku��)��
֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)������ϵ�y(t��ng)��Ӱ�
1. ֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)������ϵ�y(t��ng)�Ϳ�����Ӱ푵����ÙC(j��)�����߷���(y��ng)��Ҫ���N��͵ļ�(x��)���V��(li��n)ϵ����N�B(y��ng)��ͨ�^(gu��)�{(di��o)��(ji��)��Ϣ���ӵ�ጷŻ���M(j��n)�ܰͼ�(x��)��������̖(h��o)�Ļ��Զ�����Ҫ���á�PUFA����ͨ�^(gu��)Ӱ푼�(x��)��֮�g��(li��n)ϵ��Ӱ푄�(d��ng)������ϵ�y(t��ng)��һ���f(shu��)��(l��i)������е�֬���������N�C(j��)��Ӱ푙C(j��)�w������ϵ�y(t��ng)�Ϳ�������
��һ�N��ͨ�^(gu��)Ӱ푼�(x��)��Ĥ��֬M�ɣ��@�c(di��n)�Ͳ����������֮̎���~����е�֬����M��Ӱ�����(x��)��Ĥ��֬(Phospholipid)��֬����M�ɣ����^(gu��)��(l��i)����(du��)�C(j��)�w�ֿ������������h(yu��n)��Ӱ푣���?y��n)��S�����ߑ�(y��ng)��(y��ng)����(x��)��(Leucocyte)��(x��)��Ĥ������Þ���A(ch��)�ģ�
�ڶ��N�C(j��)����֬�����(hu��)Ӱ�����ϵ�y(t��ng)�D(zhu��n)�Q��̖(h��o)��׃�Q���@����������֬����Ӱ푵���ø
C(Protein kinase C,PKC)�Ļ�������ģ�
�����N���ÙC(j��)����֬ĽM��ͨ�^(gu��)Ӱ푏Ļ�����ϩ��(18��4n-6, Arachidonic Acid, ARA)����ʮ̼��ϩ��(20��5n-3, Eicosapentaenoic Acid, EPA)����ʮ��̼��ϩ��(22��6n-3, Docosahexaenoic Acid, DHA)�Ϳ��ܵ����� PUFA��ǰ�w���|(zh��)�D(zhu��n)���a(ch��n)�����������ԵĶ�ʮ����(Eicosanoids)���Ķ�Ӱ푙C(j��)�w�����߷���(y��ng)�Ϳ�������������DHA��ARA������Ĥ����Ҫ�M�ɳɷ֣��cĤ������(d��ng)�ԡ���Ϣ���f�����w�����������P(gu��n)��
�о���������(x��)��Ĥ֬����M���c�z�����P(gu��n)���z���֬����ɷ�׃���Ɍ�(d��o)��Ĥ�Y(ji��)��(g��u)��׃�����M(j��n)һ������Ĥ���ܵĸ�׃,�Ķ�Ӱ����ɼ�(x��)�������ߡ�����, ����(x��)����(du��)��ԭ���R(sh��)�e�����á�EPA��ARA���M(j��n)һ�������鲻ͬϵ�к;��������ܵĶ�ʮ̼�����ǰ������
(Prostaglandins, PGs)������ϩ(Leukotrienes, LTs)��Ѫ˨��,�@Щ�ɷ������װY���|(zh��)���c�C(j��)�w���߷���(y��ng)��ǰ������߀ԭø�Ͱ���ϩ�M�ɼ�(x��)������|(zh��)���ӣ��@Щ���������ٷ���ϵ�y(t��ng)��һ���֡��װY����(y��ng)�̼�ǰ�����غͰ���ϩ���γɣ�ǰ������߀ԭø�Ͱ���ϩ���γɌ����M(j��n)���ɼ�(x��)����������(x��)���_ʼ���ɼ�(x��)�����V�x�v���ʮ�������ڼ�(x��)�������wˮƽ�ό�(du��)��(y��ng)����(y��ng)���^(gu��)���Юa(ch��n)���ģ����ж�N�������ܡ�������ǰ�w��ARA����EPA�a(ch��n)�����ʮ������������Ա���ARA�a(ch��n)����������(j��ng)��(zh��ng)��������ARA�a(ch��n)�����ʮ���ᡣ��ˣ��w��(n��i)�ʮ��������þ�ȡ�Q��ARA�cEPA�ı�������ARA/EPA�����������ʮ������ԣ�����Ѫ�ܹ��ܺ��װY����(y��ng)�ȣ���֮����EPA/ARA������(hu��)�����ʮ��������á�
n-3֬�������Ը�(j��ng)��(zh��ng)��ʽӰ�ARA�Ĵ��x����׃���x�a(ch��n)�����ͣ��a(ch��n)��Ч�ܲ��ߵġ�3��ϵ��ǰ�����غ͡�5��ϵ�а���ϩ���M(j��n)���p�p�C(j��)�w�װY����(y��ng)�����o(h��)����ϵ�y(t��ng)���ܓp����n-3֬�����Ӱ푼�(x��)��Ĥ�Y(ji��)��(g��u)�������ԡ���(w��n)���Ժ�����(d��ng)��,Ӱ푼�(x��)���\(y��n)��(d��ng)�����w�γɡ����w�c���w�ĽY(ji��)�ϵȣ��Ķ��p�ټ�(x��)�����ӵĮa(ch��n)����ጷţ�n-3PUFA�ڼ�(x��)��ˮƽ�T��(d��o)�ĸ�׃�^��־ã�����Ч�������{(di��o)���I(y��ng)�B(y��ng)�أ�PUFA��(du��)ˮ�a(ch��n)��(d��ng)������ø�������(l��i)��Ҫ�Ѓɂ�(g��)���棺һ�ǾS�ּ�(x��)��Ĥ�Y(ji��)��(g��u)�͙C(j��)�ܵ������ԣ����ǘ�(g��u)�ɽy(t��ng)�Q���ʮ����ĸ���������Է����ص�ǰ�w��
EPA��DHA��ARA���c�Sϵ��(x��)��Ĥ�ĽY(ji��)��(g��u)�͙C(j��)�����P(gu��n)�����ǣ��c��������(d��ng)���෴���~�(x��)��Ĥ�еĸ߶Ȳ��֬������DHA��EPA������ARA�����^�١��~�ijЩ�M�����������ۺ���(j��ng)�M����DHA��EPA�ĺ����e�ߣ���ˣ���(du��)����У�-3PUFA����Ҫ��Ҳ�ߣ��������֬������������m�˕�(hu��)������N����(Yamamoto ��,1992)��
2.֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)������ϵ�y(t��ng)�Ϳ�������Ӱ�
Blazer��Wolke��1984�������^�쵽�����ˮ�a(ch��n)��(d��ng)������ߑ�(y��ng)��(y��ng)�Ϳ���������l(f��)�]����Ҫ�����á��~�w�M���ж�֬����(PUFA)�ĺ�������(du��)���������(d��ng)���PUFA�Ǽ�(x��)��Ĥ����Ҫ�M�ɲ��֣�����~�w�M���������ܵ�֬������ᔡ��Ӱ푡��~���r��з��ؐ��ˮ�a(ch��n)��(d��ng)�һ���ȱ��n-3PUFA��n-6PUFA����ϳ��������@Щ֬�����횏�ʳ���Ы@�ã��DZ����֬���ᡣͨ������ˮ�~�ռZ�еā�����(18��2n-6, linoleic acid , LOA)��������(18��3n-3, lanolin acid, LNA)�ɝM�����֬����(essential fatty acid, EFA)����Ҫ������ˮ�~�ռZ�Ѕs��Ҫ�L(zh��ng)�PUFA����EPA��DHA���ܝM��EFA����Ҫ(NRC��1993)��������L(zh��ng)��ȱ��EFA��EFA��ƽ�⣬һ���(hu��)��(d��o)��ˮ�a(ch��n)��(d��ng)�����L(zh��ng)���������߹��ܵ��£��ɻ��ʽ��͡����Ч���½�������ͬ�ķN�EFAȱ���Y��ƽ��ı��F(xi��n)�Dz�һ�ӵġ�
1. �����֬Ӱ푼�(x��)��Ĥ��֬�|(zh��)��֬����M�ɣ�����(x��)��Ĥ��֬����M���֛Q���˼�(x��)��Ĥ������(d��ng)�Լ�ͨ�ԣ��Ķ�Ӱ푼�(x��)�����������ܡ����յ��C(j��)�w��(n��i)PUFA����Y(ji��)��(g��u)֬�����֬��đ�A(phosphatidylcholine, PC)����֬���Ҵ���(phosphatidylethanolamine, PE)�ȣ���(g��u)����֬�p��Ĥ���Ǽ�(x��)��Ĥ�́���(x��)���Y(ji��)��(g��u)����Ҫ�M�ɳɷ֣�ֱ��Ӱ푼�(x��)��Ĥ������(d��ng)�Ժ�ͨ�ԣ�������ijЩ�I(y��ng)�B(y��ng)���|(zh��)(��֬��)�����������գ����c����֬ɷ�(��đ�̴�)�������c�\(y��n)ݔ���M(j��n)��Ӱ��~����L(zh��ng)�l(f��)�����rз����w��͑Ƥ�c׃�B(t��i)�������֬���|(zh��)������(hu��)Ӱ푺��(d��ng)���ǰ���ٺϳɡ��wҺ���߷���(y��ng)�Լ����ɼ�(x��)�����������ã�Sheldon��Blazer(1991)Ҳ�C�����в�ͬ��֬����ϣ���Ӱ푰��c(di��n)��β�t(Ictalurus punctatus)���ɼ�(x��)���Ě������ԣ����Қ��������c������L(zh��ng)朸߶Ȳ��֬���Ậ���������P(gu��n)��
2. ֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)�������Ӱ푡�ȱ������֬����r(sh��)�����ɼ�(x��)�����缚(x��)���������ͣ����w�Įa(ch��n)���p�١�Kiron�ȣ�1995a���о��l(f��)�F(xi��n)����(d��ng)���V�~(Oncorhynchus mykiss)�ι����PUFA�ռZ���~��и��õĵֿ���(x��)����Ⱦ��������������ռZ��n-3PUFA����������~�����������Montero��(1998)��(b��o)�����ռZ�к�������n-3PUFA��(du��)�ھS�ֽ��^��(Sparus aurata)�Ⱥ����~��a(b��)�w������������ġ�ͬ�r(sh��)ֵ��ע����ǣ��ιn-3/n-6֬��������ߵ��ռZ���Է�ֹ�������q(Salmo salar)���ʲ��İl(f��)���������䌦(du��)��(y��ng)����������(Bell�ȣ�1991)����ˣ�n-3֬����H��(du��)�ھS���~�������L(zh��ng)�����Ч�ʶ����DZ���֬���ᣬ���Ҍ�(du��)�����������Ч�ʺ���Ѫ�ܙC(j��)�ܶ���Ҳ�DZ���֬���ᡣ���⣬�N���ﻯ�W(xu��)�Į�������(y��ng)�ͼ������c�ι��PUFA�����ᔡ���ռZ������֬��������(d��o)�µ����I(y��ng)�B(y��ng)��?f��)pʧ���P(gu��n)���B(y��ng)ֳ��(ch��ng)�г�Ҋ�����p����Ƥ�w�l(f��)�׆�(w��n)�}Ҳ�c֬��������P(gu��n)(Lall�ȣ�1993��Olivier��1993)��
3. ��ͬ��֬����M�Ɍ�(du��)ˮ�a(ch��n)��(d��ng)������(x��)�������߷��ӵ�Ӱ푲�ͬ��һ����r�ǣ�n-3PUFA��(du��)�C(j��)�w���߾����������ã���n-6PUFA��(du��)�C(j��)�w���߾��д��M(j��n)���á��~�ͺ�ֲ�����в�ͬˮƽn-3��n-6֬���Ậ���ɸ�׃���Ŀ�~(Paralichthys olivaceus)�ʹ������q��(x��)����֬��֬����ĽM��(Bell�ȣ�1991a��1993��1994)����֬��֬����M�ɵ�׃����(hu��)Ӱ��ʮ����ǰ�w��ĺϳɡ�
���(n��i)���ɵ�(1992)�ò�ͬ�����֬(�������������������������x֬����)��(qi��ng)�����u�xͶι�o�lʯ��(Oplegnathus fasciatus)������(Pagrosomus major)���Ա��^�䌦(du��)�~����L(zh��ng)���������������D��Ч������(sh��)�(y��n)�Y(ji��)���@ʾ���~�w��HUFA�����c�u�x��PUFA���������õ���P(gu��n)ϵ����PUFA�zȡ���õ����x֬����Mֻ�õ��͵Ļ�������֮?d��ng)zȡHUFA����(y��u)�|(zh��)��������֬���e�ǽo�����~Ͷι��������(qi��ng)�����u�x�M���@���˃�(y��u)��������ɿ�(j��)��
֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)������ϵ�y(t��ng)��Ӱ�
4. ֬���ጦ(du��)�~�(du��)���ɼ�(x��)���ۺ��w(Macrophage aggregates, MAS)��Ӱ푡��������~�Ƣ���I����е�MAS�ж�N�������ܣ����缚(x��)���U�����Դ�����|(zh��)����ѭ�h(hu��n)/��(ch��)��/�ⶾ������?c��)��خ������߷��?y��ng)��Ҳ�l(f��)�]����Ҫ�����ã����ҿ������齡��ָʾ��(health indicators)��MAS���ܶ�(density)����С(size)��ɫ�غ���(pigment content)��׃���L(zh��ng)����Ⱦ�O(ji��n)�y(c��)��ָʾ����Ȼ��MAS�Ĕ�(sh��)���ͽY(ji��)��(g��u)Ҳ��(hu��)�ܵ��������ص�Ӱ푣�����һ�㑪(y��ng)�����~ĠI(y��ng)�B(y��ng)��r�ȡ�Montero ��(1999)�о��l(f��)�F(xi��n)�������ȱ��n-3HUFA�܉����ӽ��^��Ƣ�K�е�MAS�Ĕ�(sh��)�����Ķ�����(qi��ng)�C(j��)�w������������
֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)������ϵ�y(t��ng)��Ӱ�
5. �D����EPA/DHA��(du��)�~����L(zh��ng)��Ӱ푡��о������������~�D����EPA/DHA�^(gu��)�ߣ���(du��)�~����L(zh��ng)����������ԭ����EPA��ֹ��ARA����ǰ������(PGs)���M��ARA��PGs��ǰ�w,��һ���J(r��n)��EPA��ARA��������Ҫ(Sargent et al,1993��1995a)������m�˱�����EPA�cARAҲ�ܵ��ձ��P(gu��n)ע��
DHA�����u���������ǵ�ˮ�q�~�w����Ҫ��ǰ�����أ������ƺ�ˮ�����~��EFAȱ������������L(zh��ng)�������ɻ��ʽ��͵Ƞ�r���F(xi��n)���C��(sh��)(Rainuzzo��, 1994; Rodriguez��,1994), EPA�����յ��εȽM��������Ĥ��,�M����δ̽��EPA���@Щ�M���е����á����⣬EPA߀��E3����-E3��ǰ�����ص�ǰ�w���|(zh��),��߀�cARA��(j��ng)��(zh��ng)������ARA�D(zhu��n)����ǰ�����ء��N��ˮ�����~�����I���g��ARAҲ��(hu��)�����x��ر�����(l��i)���@�f(shu��)��ԓn-6ϵ�еĸ߶Ȳ��֬����Ҳ����Ҫ�ı���֬���ᡣ�mȻ�������~��һЩ�M����ARA�ĺ�������EPA�ĺ�����������E2����-E2��ǰ�����ص���Ҫ��(l��i)Դ�����ں�ˮ�����~����֬�����������^�ߵĺ����������(l��i)���о�߀�C��(sh��)��ARA����Ҫ�ԣ��M��ԓ֬�������DHA��EPA�ǘ���ߺ�ˮ�����~�����L(zh��ng)�ٶȣ������ƺ���(du��)��ߺ�ˮ�����~�ijɻ�������Ҫ����(Castell,1994)��
�D���� EPA�����^(gu��)�߶� DHA�����^(gu��)�ͣ����ܕ�(hu��)��(d��o)������Ĥ��֬��DHA��EPA�����IJ�ƽ�⡣Watanabe(1993 )�J(r��n)�飬�����~����Ĥ�� DHA�c EPA������ʧ����ܕ�(hu��)���@���������~�����ⲿ�����������ԣ��M�� EPA���܌�(du��)����Ĥ�Y(ji��)��(g��u)�a(ch��n)������Ӱ푵ęC(j��)���в������
Bell��Colleagues(1993 )�о��������ι���Ӂ�����(�����Ậ����)��ϵĴ������q�İ�(x��)����֬�е�EPA�������ι�����~�ͺ����տ�������ϵ�Ҫ�ߣ�������иߺ����ā�������Ԝp��?g��u)�ARA������PGE2��TXB2�Įa(ch��n)�����@�NЧ��(y��ng)�ĽY(ji��)��������(qi��ng)�˙C(j��)�w�Ŀ��l(f��)�������͜p�p���K�ēp����Bell��Colleagues(1994)��(du��)������(Scophthalmus
maximus)���~�_չ��ƵČ�(sh��)�(y��n)�����õ������ƵĽY(ji��)�����ι���Ӂ�������ϵ��~M���е�EPA�����^�ߺ�ARA�^�ͣ������S��TXB2��PGE�������͡� ֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)������ϵ�y(t��ng)��Ӱ�
Bell��Colleagues(1996)�l(f��)�F(xi��n)���ι����зքe�������տ�����(n-3PUHA������)�́�����(n-3PUFA������)�Ĵ������q��s�a(ch��n)����ͬ����EPA�������ɴ˵ó�n-3PUHA�ܸ��õ����ƏĶ�ʮ�����D(zhu��n)����n-6PUFA�����c(di��n)��β�t����иߺ���n-3PUHA��(hu��)�����伲���ĵֿ���(Fraclossi��Lovell,1994)�����ɼ���������(Lingenfelser��,1995)��Li��Colleagues�J(r��n)�飬�ߺ�����n-3PUHA�����~����ߙC(j��)�ܵĿ��ܙC(j��)���c��������ƣ���Ҫ�ǽ������ɼ�(x��)�������Լ�(x��)����LTB4�Įa(ch��n)���Լ���������LTB5�Įa(ch��n)����Fraclossi��Lovell(1994)�J(r��n)�飬���c(di��n)��β�t�����n-3PUHA��n-6PUHA�����m�˵ı�����������������߹��ܡ�Tatner(1996)��Henderson(1996)�^�쵽��Ͷι��n-3PUHA/n-6PUHA��ֵ��ϵĴ������q�ڸ�ȾAeromonas salmonicida �cVibrio anguillarum��ԭ�w�r(sh��)��������(qi��ng)��B�ܰͼ�(x��)���ķ���(y��ng)����ߴ���ʡ�Ashton��Colleagues(1994)�о��������ι����n-3ϵ��֬������ϵĺ��V�^�I��{�Ҹ�Һ���D(zhu��n)�ƴ̼��������ι����n-6ϵ��֬������ϽM�ď�(qi��ng)��
�M��Ŀǰn-3PUFA��(du��)�~����߷���(y��ng)Ӱ푵Ĉ�(b��o)������һ�£����Ǹ���n-3֬������ռZ�_��(sh��)�������(x��)��Ĥ�ķ�(w��n)����(Erdal�ȣ�1991��Klinger�ȣ�1996)���@��(du��)��ˮ�a(ch��n)��(d��ng)���(l��i)�f(shu��)�@���Ȟ���Ҫ����?y��n)������ڜض�׃������ˮ��Ҫ�S�ּ�(x��)��Ĥ�����Ĺ�����Ҫn-3֬���ᡣ
6. �ضȌ�(du��)֬�������~����߷���(y��ng)�е�Ӱ푡�֬���ጦ(du��)�~����߷���(y��ng)��Ӱ��c����̎�ĭh(hu��n)���l���������P(gu��n)����ضȾ͕�(hu��)Ӱ�֬���ጦ(du��)�~����߷���(y��ng)��Ч��(y��ng)��Lingenfelser��(1995)�о��l(f��)�F(xi��n)���ڵؗ͜l���£����c(di��n)��β�t�����n-3ϵ��֬���Ậ���ߕr(sh��)�͕�(hu��)����(qi��ng)�����߹���(�e������(qi��ng)��(x��)������������),���ڸߜؗl���£��ι���c(di��n)��β�tn-6ϵ��֬���Ậ���ߵ���ϕr(sh��)��������(qi��ng)�伲���ֿ�������ˮ�ص��@�N��������Ҳ�S�cn-3֬����S�ּ�(x��)��Ĥ����(d��ng)�ԵĹ������P(gu��n)�����������ɾ����õĔzȡ��B(t��i)�r(sh��)�������
֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)������ϵ�y(t��ng)��Ӱ�
Fraclossi �� Lovell(1994)Ҳ��(b��o)���˲�ͬ�Ĝضȗl����(hu��)Ӱ�֬���ጦ(du��)���c(di��n)��β�t������Ч��(y��ng)���ض�Ӱ푲�ͬ֬���ጦ(du��)�~����߷���(y��ng)��Ч��(y��ng)������������n-3ϵ��֬������Ҫ�ǾS�ּ�(x��)��Ĥ������(d��ng)�ԣ��@�c(di��n)��(du��)���ɼ�(x��)���Ĕzȡ�dz���Ҫ��n-3PUHA�ձ鱻�J(r��n)���ڵؗ͜l���������Ӽ�(x��)��Ĥ������(d��ng)�ԡ����g�Ժ�ͨ�ԣ�����(du��)����ˮ�a(ch��n)��(d��ng)���(l��i)�v���DZ���֬���ᣬ����ͬˮ�a(ch��n)��(d��ng)��N��ڲ�ͬ�h(hu��n)���l���£���(du��)����֬�����������һ���IJ���@�N����������ڲ�ͬˮ�a(ch��n)��(d��ng)�ﱾ��֬�Ҳ������Ҫ�Ǽ�(x��)��Ĥ��֬֬����M�ɵ��خ��ԛQ���ġ�֬����Ӱ푙C(j��)�w�����߷���(y��ng)�����_(d��)�̶���ȡ�Q���ڼ�(x��)��Ĥ�϶�ʮ�����ǰ�w���|(zh��)������к��и�ˮƽ��n-6PUHA��������(qi��ng)���߷���(y��ng)�����ځ�(l��i)��ARA�ľ��м������õĶ�ʮ���������ӣ�������к��и�ˮƽ��n-3PUHA�t�������߷���(y��ng)�����ځ�(l��i)��EPA���l(f��)�Ķ�ʮ����a(ch��n)�����ӡ�
Ҳ���C��(j��)����������е�n-3ϵ��֬�������~�w���������ߴ̼�����Ч����
Ŀǰ�˂���(du��)ˮ�a(ch��n)��(d��ng)�������ϵ�y(t��ng)߀δ��ȫ�о�������e��, ˮ���o(w��)����(d��ng)�����ߙC(j��)�ƵĻ��A(ch��)�о�߀���^�t��������һЩ�u(p��ng)�r(ji��)��(bi��o)��(zh��n)���z�y(c��)ָ��(bi��o)�y�Դ_�����z�y(c��)�ֶ�����ԭ���Ƽs��ˮ�a(ch��n)��(d��ng)�����ߌW(xu��)�İl(f��)չ��ͬ�r(sh��)Ҳ�Ƽs��
���(y��ng)���b�������P(gu��n)�W(xu��)�Ƶļ��g(sh��)���������ֶΣ��_չ֬���ጦ(du��)ˮ�a(ch��n)��(d��ng)������ϵ�y(t��ng)���ÙC(j��)�Ƶ������о�������һ���m��ˮ�a(ch��n)��(d��ng)��֬����I(y��ng)�B(y��ng)�c�����߹��ܼ����������о��͙z�y(c��)�u(p��ng)�r(ji��)�ķ������M(j��n)һ���о�ʹ�ò�ͬ֬��Դ��(du��)ˮ�a(ch��n)��(d��ng)�����߹��ܺͿ�������Ӱ푣��L(zh��ng)���M(j��n)��ˮ�a(ch��n)��(d��ng)�����߹��ܺͼ������εľC���{(di��o)���о����ɞ��B(y��ng)ֳˮ�a(ch��n)��(d��ng)����m(x��)׃�Q֬��Դ�ṩ��Ҫ���Y�ϡ�
ˮ�a(ch��n)��(d��ng)��������ӄ���ԇ�}
1.��(d��ng)ǰˮ�a(ch��n)��(d��ng)�����ø�Ƅ��о��_�l(f��)�c��(y��ng)���д���ʲô��Ҫ��(w��n)�}�����(y��ng)
����_�l(f��)ˮ�a(ch��n)���ø�Ƅ���
2.Ո(q��ng)���U��һ��Ŀǰ���еĎN���B(t��i)�Ƅ������ÙC(j��)�������Y(ji��)��ˮ�a(ch��n)��(d��ng)�P
ˮ�a(ch��n)�B(y��ng)ֳ�����c(di��n)��Փ��ˮ�a(ch��n)��(d��ng)��������B(t��i)�Ƅ��о��_�l(f��)��˼·����ˇ�^(gu��)
�̣�
3.Ӱ�ˮ�a(ch��n)��(d��ng)����ɫ��ʹ��Ч����������Ҫ����Щ���_�l(f��)��Ч��ˮ�a(ch��n)��(d��ng)����
ɫ����(y��ng)ע����Щ�P(gu��n)�I�ԵĆ�(w��n)�}��
4. Ո(q��ng)�U���S����C Ӱ�ˮ�a(ch��n)��(d��ng)���������Ŀ��ܙC(j��)�ƣ���ԇ��Ӱ푾S����C��
��ˮ�a(ch��n)��(d��ng)�����ߙC(j��)�ܵ����أ�
5.�Y(ji��)�Ϯ�(d��ng)ǰˮ�a(ch��n)��(d��ng)��������ӄ��о��_�l(f��)����(y��ng)�ìF(xi��n)��U�����ˮ�a(ch��n)��(d��ng)���
�����ӄ��о������c(di��n)���y�c(di��n)�c���c(di��n)��(w��n)�}��
|
|


















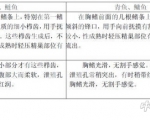

















 ���п�
���п�
 �l(f��)���� 2011-6-23 09:55:04
�l(f��)���� 2011-6-23 09:55:04

 �D(zhu��n)��
�D(zhu��n)�� ����
���� ����
���� �ղ�
�ղ� ֧��
֧�� ����(du��)
����(du��) ������
������ ��픿�
��픿� ׃ɫ��
׃ɫ�� ǧ���
ǧ��� �@����
�@���� �l(f��)���� 2011-6-23 12:30:12
�l(f��)���� 2011-6-23 12:30:12





